인지 신경과학
서 유 헌 (서울대학교 의과대학 약리학 교실)
인지 과학 : 김영정, 김재권, 조인래, 김정오, 조명한, 이정민, 김영주, 김영택, 장병탁, 김형주, 백은옥, 강봉균, 서유헌, 이경민, 최재천 공저, 태학사, 2000. Page 421~480
(1) 흥분이 일어나지 않는 안정시에 나타나는 신경세포막의 막전위 (안정전위)
2) 학습과 기억 (Learning and Memory) 의 정의
3) 신경세포접촉 (Cell assemblies) 과 시냅스
6) 관계학습 (Associative Learning)
7) 능동적 조건 반응 (Operant Conditioning)
(3) 관찰 학습 (Observational or Vicarous Learning)
(1) 장기강화 (Long-term Petentiation)
(2) 학습과 기억의 약리 (Pharmacology of Learning and Pharmacology)
신경계는 중추신경계와 말초신경계로 구성되어 있다. 인체의 모든 조직과 기관들은 중추신경계 (中樞神經系) 에 의하여 기능이 조절되고 통괄됨으로써 하나의 통일된 개체로 활용할 수 있다. 중추신경계에 있는 신경세포의 긴 가지들이 신체 각 부분으로 나가서 말초신경계 (末哨神經系) 를 구성하고 있다. 포유동물 중추신경계는 원래 관 모양의 관상 구조로부터 발달하여 뇌 (腦) 와 척수 (脊髓) 가 되었다. 관 모양의 구조는 뇌에서는 뇌실 (腦室) 이 되고 척수에서는 중심관 (中心管) 이 되었다. 이 속에 뇌척수액이라고 불려지는 물로 차있으며 이 척수액은 뇌 실에 있는 맥락총에서 생산되어 뇌실계를 돌아 뇌간에서 표면으로 나와 뇌를 감싸 돈 다음 정맥으로 흡수된다. 이 뇌척수액은 뇌를 감싸 중추신경계를 띄움으로써 뇌에 대한 외부로부터의 기계적인 충격을 완충해주며 뇌에서 생산된 대사물을 제거해준다. 뇌 속을 관류하고 있는 뇌척수액은 약 100cc 정도 되며 하루에 약 500cc 정도 생산되고 있기 때문에 하루에 5번 정도 교체되고 있다. 뇌는 앞쪽에 있는 전뇌 (前腦), 중간부위에 있는 중뇌 (中腦) 와 뒤쪽에 있는 후뇌 (後腦) 로 구성되어 있다. 전뇌는 뇌의 가장 바깥껍질을 형성하고 있는 대뇌피질 (大腦皮質) 과 변연계 (邊緣系) 로 구성되어 있다. 후뇌는 생명중추 (혈압중추, 호흡중추 등) 가 있는 뇌의 밑뿌리에 위치하고 있는 연수(延髓; medulla) 와 뇌교 (腦橋) 그리고 평형과 운동을 주관하고 있는 소뇌 (小腦; cerebellum) 가 속하고 있다.
변연계는 뇌의 중심부 아래에 자리하며 계통 발생상 뇌에서 비교적 일찍 발생된 부분으로서 고피질 (古皮質) 이라고 불려지고 있으며 늦게 발달한 신피질 (新皮質) 인 대뇌피질에 의해 완전히 둘러싸여 있다. 대뇌피질은 외부 환경과의 교신을 통하여 이를 입체적으로 인식하는 능력을 가지며 목적 지향성 이성행동을 주재한다. 즉 고도의 사색기능, 판단기능, 창조적 정신기능 등의 고등정신활동을 하는 곳이며 운동과 감각을 또한 주재하는 곳이다. 이에 비하여 번연계는 본능 행동과 정서 감정을 주재하는 기구로서 행동의 의욕 (意慾), 학습 (學習), 기억 (記憶) 과정에 깊이 관여한다.
대뇌피질의 표면은 신경세포체가 밀집되어 있기 때문에 회색으로 보이는 회백질 (恢白質) 부외와 속 부분은 흰 신경섬유로 이루어져 있기 때문에 희게 보이는 흰 백질 (white matter) 부로 구성되어 있다. 대뇌를 외관상 보면 꼬불꼬불한 고랑과 둑이 많이 보인다. 휘어진 고랑을 구 (溝; sulcus), 깊은 고랑을 열 (烈; fissure), 둑을 회전 (回轉; gyrus) 이라 부른다. 좌우 대뇌반구는 앞쪽의 전두엽 (前頭葉), 위쪽의 두정엽 (頭頂葉; parietal lobe), 뒤쪽의 후두엽 (後頭葉; occipital lobe), 뇌 가운데 있는 변연엽 (limbic lobe) 의 다섯 가지 엽으로 나누어 진다. 대뇌피질은 기능과 구조적 특징에 따라서 브로드만 (Brodmann) 이 1번에서 52번까지 번호를 붙여서 세분하였다.
우리의 두뇌는 좌 우측 2개 대뇌반구를 갖고 있다. 뇌 반구는 우리의 사고를 관장하고 신체의 기능을 조절한다. 뇌의 좌측 반구는 몸의 오른쪽을 조절하고 우측 반구는 몸의 좌측을 담당한다. 뇌의 중간에서 나뉜 이유에 대한 하나의 가설은 만일 한쪽이 손상을 받으면 나머지 하나가 보완한다는 것이다. 이런 이원성은 다른 내부기관 즉 폐, 신장 그리고 눈, 귀, 사지 등에서 흔하게 관찰된다.
출생 시 뇌가 손상 받았을 때는 종종 건강한 대뇌반구가 손상된 뇌의 기능을 성공적으로 보완한다. 수술로 한쪽 반구가 제거된 영아에게 육체적, 정신적 성장이 거의 정상적으로 이루어지는 것을 볼 수 있다. 성인에게는 영구적인 언어 상실을 초래할 수 있는 뇌졸증을 일으킨 6~7세 어린이는 심각한 언어 발달 장애를 겪지 않는 경우가 흔하다. 그러므로 아이들이 나이가 들수록 적응력은 감소하고 대뇌반구가 전문화되는 것이 정상적이다. 비록 우리의 뇌가 두쪽으로 나뉘어 있다 해도 두 반구는 동일하지는 않다. 좌우 대뇌 반구는 뇌략 (腦樑, Corpus Callosum) 이라고 불리는 큰 다리에 의해서 연결되어 있다. 간질을 치료하기 위해 환자의 두 반구를 연결하는 뇌량을 절단하는 치료 방법을 사용하였다. 간질 발작이 일어날 때 뇌의 신경세포들은 때때로 "전기폭풍" 이라고 묘사되는 비정상적이고 불규칙한 전류를 뇌량에 방전한다.
신경외과 의사들은 뇌량을 절단함으로써 이 폭풍을 진정시킬 수 있다. 뇌량의 절단은 치유 불가능한 간질의 보편적인 치료법이 되었다. 머리 안에 두 개의 뇌가 존재한다는 말도 일리가 있다. 이것은 각각의 대뇌반구가 독립적으로 작동할 수 있으며 한쪽 뇌반구만으로도 살 수 있다는 것을 의미한다.
비록 분리된 대뇌반구가 기능은 잘하나, 뇌의 좌우반구가 분리된 환자는 영상과 언어를 연결시키는 기능에 상당한 장애를 가질 수 있다는 사실이 그후 연구에서 밝혀졌다. 두 대뇌반구는 독립적으로 가능한 것이 아니라 상호 작용하는 것이다. 즉, 한 손만으로도 일을 할 수 있으나 두 손을 쓸 때보다 훨씬 불편한 것과 비슷하다.
대뇌피질은 대뇌반구 표면의 회백질이며, 두께 평균 2.5㎜, 표면적은 2,300㎠ 가량으로 신문지 한 장 넓이이다. 용적은 550㎤ 가량으로서 그 속에 포함되는 뉴론 수는 대략 150억으로 추정된다. 대뇌피질은 편의상 가장 앞쪽에 있는 전두엽 (前頭葉), 머리 꼭대기에 있는 두정엽 (頭頂曄), 머리 뒤쪽에 있는 후두엽 (後頭葉), 머리 옆쪽에 있는 측두엽 (側頭葉) 으로 크게 나눈다. 이밖에 대뇌 반구 속에는 먹는 배모양의 이상엽 (梨狀葉) 그리고 뇌척수액이 돌아다니는 측뇌실 (側腦室) 의 밑변에는 해마 모양을 한 해마 (海馬; hippocampus) 등을 통틀어 변연엽(邊緣葉) 이라 부르고 있다.
대뇌피질은 여서 층의 세포로 구성되어 있는데, 다만 계통발생상 오래된 변연엽을 고피질 (古皮質) 이라 하고, 나머지 대뇌피질은 비교적 늦게 발달되었기 때문에 신피질 (新皮質) 이라 한다.
신피질에서 볼 수 있는 주요한 뉴런은 원추모양의 추체 (錐體) 세포와 과립모양의 과립세포를 가지며 그 첨단부에서 피질표면으로 향하여 첨단수상돌기가 나오고, 기저부에서 피질하로 축삭돌기가 뻗어 있다.
이와 같이 대뇌피질은 수평으로 나란히 배열한 세포층이 중첩된 구조이지만 뉴런을 연결하는 축삭돌기의 방향은 수직으로 뻗어 있어 수평으로의 뉴런 연결을 제1층 분자층을 제외하면 미약하다. 따라서 흥분의 왕래는 주로 수직방향이며 수평으로의 흥분전도는 드물다. 그래서 대뇌피질의 기능 단위는 수직방향으로 연결된 뉴런기둥이라 하겠다.
아래에서 올라온 흥분을 대뇌피질부에서 최종 집합하여 명령을 수직방향으로 내려보내 적절한 행동을 하도록 하고 있다.
우리 몸의 감각을 관찰하는 중추 부위는 브로드만 영역 1, 2, 3이고, 미각은 영역 43, 시각은 영역 17, 청각은 영역 41, 42 이며 운동을 관할하는 뇌 중추부위는 영역 4,6이며 언어영역은 영역 44,45이다. 주변부에 있는 연합영역이 위와 같은 고등기능을 보조하고 있다.
<신피질의 분류>
감각영역
체성감각영역: 영역 1, 2, 3
미각영역 : 영역 43
전정감각영역 : 영역 2
시각영역 : 영역 17
청각영역 : 영역41, 42
감각연합영역
체성감각연합영역: 영역 5, 7
시각연합영역 : 18, 19
고등시각연합영역: 영역 20, 21
청각연합영역 : 영역 22
운동영역
영역 4, 6
보조운동영역 : 영역 6
전두안영역 : 영역 8
운동성언어영역 : 영역 44, 45
고등연합영역
전전두영역 : 영역 9, 10, 11, 12
두정엽 후하부 연합영역 : 영역 39,40
수의적 근육운동에 관여하는 운동영역은 전두엽과 두정엽을 분리하는 중앙고랑인 중심구 (中心溝) 의 바로 앞에 있다. 근운동에 영향을 미치는 피질하 구조로 뇌의 가운데에 기저핵 (基低核) 이 있다. 소뇌는 위에서 말한 운동피질, 기저핵과 긴밀한 연락을 가지며 근운동의 정밀 조정 역할을 한다.
운동영역은 신체의 반대측 반신이 표시되어 있으며 신체상부의 근육은 운동영역 아랫부분이 지배하고 신체 하부의 근육은 운동영역 윗부분이 지배한다. 즉 상하 좌우가 뒤바뀌어 있다.
운동피질의 면적은 근육의 크기보다 운동의 정밀도와 복잡성에 따라 결정된다. 즉 몸통보다 손, 입, 혀 등 세밀하고 정교한 운동에 관여하는 부위가 특히 넓은 자리를 차지한다. 이는 인간이 손을 놀리며 일하고 입으로 말하며 생활하기 때문이다. 인간 생활에서 근운동은 입을 중심으로 한 말하기와 손을 중심을 한 작업이 주이기 때문에 이곳을 지배하는 대뇌피질의 면적이 넓어진 것이다.
한쪽의 운동영역이 파괴되면 반대측의 근 마비가 오게 된다. 감각영역은 운동영역 중앙고랑인 중심구 뒤쪽, 즉 바로 뒤에 있는 후중심회전(後中心回轉) 부위에 있다. 운동영역과 마찬가지로 감각영역도 감각이 정밀한 손, 입등은 넓게 표시되고 감각이 둔한 몸통부위는 좁게 표시되어 있다. 중풍 시에 한쪽 뇌혈관이 파열되거나 뇌혈관이 막히면 반대측 근육운동이 마비되거나 감각이 마비된다. 어느 부위가 침범되느냐 따라 운동과 감각마비가 결정된다. 양쪽 운동과 감각 영역 모두가 침범되면 전신 불구가 되며 한쪽이 전부 침범되면 반신불수가 될 수 있는 것이다. 그러나 극히 일부분이 침범되면 마비증세가 미약하여 잘못 느낄 수도 있다.
청각영역은 측두엽에 위치하고 있다. 한쪽 청각영역의 장애가 있어도 난청이 오지 않는 것은 양측 청각영역이 연결되어 있기 때문이다. 시각영역은 머리 뒤쪽의 후두엽에 있다. 망막에서 시작된 시각 구심흥분은 시신경과 시각을 거쳐 몇 차례 뉴런을 교체하여 시각영역에 들어온다. 시각전도로가 파괴되면 소경이 된다. 머리 뒤쪽을 심하게 다치면 물체를 볼 수 있는 전도로는 손상을 받지 않으나 물체를 종합적으로 의식하여 식별할 수 있는 능력이 없어지게 된다. 대뇌피질에는 기이하게도 통각의 느낌과 연관된 영역이 없다. 대뇌피질은 어떤 영역을 자극하거나 절단 혹은 절제하여도 아무런 통증을 느끼지 않는다. 통각은 일차적으로 뇌의 한가운데에 있는 뇌의 미분화된 영역인 시상 (視床)에서 느껴지며 여기서 다듬어지지 않은 막연한 아픔이 의식에 떠오르면 대뇌피질이 아픔의 소재를 밝혀주는 듯이 보인다.
대뇌피질의 운동영역과 감각영역을 제외한 나머지 넓은 피질영역은 정신 기능 즉 인식, 판단, 사고, 기억, 학습 등 고등한 정신 기능이 이루어지고 있다. 그러나 대뇌피질의 모든 영역을 그 기능과 결부시켜 말하기는 아직 요원하다.
운동영역에 있는 신경세포에서는 의식적 근운동의 명령이 시발된다고 하여 흔히 운동영역을 운동중추라 한다. 그러나 사실 운동영역만으로 수의운동이 이루어질 수는 없다. 운동영역 신경세포는 아래로부터 흥분을 받아 아래로 흥분을 전달할 뿐 수의운동이 여기서 시작된다고 볼 수 없다. 즉 수의적 흥분을 아래로 전달하는 1차 운동중추의 구실을 할 뿐이다.
이 관계는 감각영역에서도 마찬가지로 적용된다. 1차 감각영역은 감각을 의식하는데 직접 관여하지는 않는다. 구심흥분을 더욱 고위 감각중추에 전달하기 위한 중계소 구실을 한다. 그러므로 의식적 지각과 이에 대한 이해, 인식을 담당하는 뇌 영역은 따로 있다고 보아야 한다. 바로 이들 수의운동의 구성과 감각의 인식기능을 담당하는 부위가 연합영역 (聯合嶺域; Association Area) 에 있으리라 믿어진다. 앞서 말한 운동영역, 감각영역에 속하는 부분을 제외한 대뇌피질에는 아직도 넓은 영역이 남아있는데 이 영역이 바로 연합영역으로서 동물이 발달할수록 연합영역 면적이 넓어지게 된다. 즉 연합영역의 발달정도가 고등 뇌 기능의 발달정도와 비례하며 문화 발달 정도를 결정한다고 할 수 있다. 인간보다 연합영역이 더 발달한 동물이 있다면 인간보다 우위에 서서 우주를 지배하게 될 것이다.
특정 뇌 부위가 손상되면 감각기능은 정상이지만 감각 경험을 분석하고 다른 감각 경험과 비교 검토하여 그 내용과 의의를 파악하는 능력이 상실된다. 인식능력이 없어진다고 하여 이를 실인증 (agnosia) 이라고 한다. 머리 뒤쪽에 있는 뇌 부위인 후두엽에는 일차 시각 영역이 포함되어 있다. 이 일차 시각영역 부위가 손상되면 봉사가 되지만 주변부의 연상영역이 손상되면 시각 기능은 이상이 없지만 보는 대상의 의미를 해독하지 못한다. 음식을 보고서 무슨 음식인지 알지 못하고 뱀을 보고도 무서움과 징그러움을 느끼지 못하기 때문에 두려움 없이 만져본다. 대상을 볼 수 있으나 이 대상의 정확한 실체를 파악하지 못하는 것이다. 이러한 것을 시각성 실인증 (視覺性失認症) 이라고 부른다.
머리 옆쪽에 있는 측두부 (側頭部) 에는 일차 청각영역이 있다. 이 일차 청각영역의 주변부 연상영역이 손상되면 소리는 들을 수 있지만 소리의 의미를 이해할 수 있는 능력이 없어진다. 이를 청각성 실인증이라 한다. 또한 청각은 이상이 없지만 대상을 만져보고 인식할 수 있는 능력이 없어지는 경우를 축각성 (觸覺性) 실인증 혹은 입체 실인증이라 부르고 있다. 이 병은 머리 꼭대기에 있는 두정엽 (頭頂葉) 부위가 손상될 때 나타난다. 특히 맨 위쪽 두뇌 꼭대기 중앙부에 있는 감각 영역 뒤의 두정부가 손상될 때는 자기의 손이 나 좌우를 구별 못하는 경우도 나타난다. 이런 경우를 자가 국소실인증 (自家局所失認症) 이라고 부른다.
이와 같이 우리 뇌는 각 부위마다 독특한 기능을 가지고 있기 때문에 이들 뇌의 손상은 심각한 신체기능장애로 나타날 수 있다.
뇌의 각성상태를 조절하는 부위이다. 특히 뇌의 가장 바깥쪽에 있는 피질 (皮質) 부위가 고도의 정신기능을 유지하는데 가장 중요하다. 뇌의 활동수준과 긴장도를 조절하는 부위를 총칭하여 망상활성화계 (網牀活性化系; reticular activating system) 라 부르며 뇌의 밑뿌리에 있는 뇌교와 연수부위에 있으며 신경섬유가 거미줄처럼 얽혀 있기 때문에 망상체 (網牀體) 라 부른다. 이 부위에서 끊임없이 상부에 있는 뇌로 흥분전파를 발사하여 뇌피질 신경세포를 끊임없이 깨어 있게 만들기 때문에 잠자지 않을 때 의식을 명료하게 유지할 수 있게 되는 것이다. 만약 이 부위가 손상되면 뇌 활동의 안정성이 깨어져서 잠든 듯 깨어난 듯하기를 되풀이하여 기억의 혼란도 오게 된다. 즉 정상적 감각흥분에 의한 각성반응은 없어지고 느린 뇌파가 나타나며 혼수상태에 빠지게 된다. 따라서 각성상태가 유지되려면 망상체가 필요하다. 수술할 때 전신마취제를 투여하면 제일 먼저 이 망상활성화계가 마취되어 외부로부터의 모든 자극에 대하여 대뇌는 인식할 수가 없게 되어 의식이 소실된다. 뇌피질에는 항상 방대한 양의 정보가 들어오고 있지만 우선 중요한 정보만을 골라내고 나머지 정보는 대개 무시해 버린다. 대뇌피질 신경세포는 강한 자극, 중요한 자극에 대해서는 뚜렷한 반응을 보이지만 중요하지 않은 약한 자극에 대해서는 반응을 보이지 않는다. 그러나 망상활성화계가 손상이 되면 대뇌피질 신경세포의 긴장도가 없어져서 자극을 구별하지 못하게 되어 강한 자극이나 약한 자극이나 비슷하게 반응한다. 즉 자극에 대한 분별력의 소실로 행동이상도 나타날 수 있다. 이와 같이 생명중추가 있는 뇌간부위에 위치하고 있는 망상활성계는 인간의 의식을 명료하게 유지시켜서 인간의 모든 활동 즉 문화창조를 가능케 해주는 의식중추이다.
호르몬을 분비하는 뇌하수체 바로 위에 있는 뇌 부위가 시상하부 (視床下部) 이다. 이 시상하부는 뇌하수체 호르몬의 분비를 조절할 뿐만 아니라 체온을 조절하는 중추이다. 이곳에는 혈액의 온도를 느끼는 감각기가 있어서 체온을 올리기도 하고 내리기도 한다.
춥고 더울 때 피부에 있는 온도 감각기가 작동하여 이 정보를 뇌에 있는 시상하부에 전해주어 체온을 조절한다. 즉, 추울 때는 우리의 의사와는 관계없이 시상하부의 명령에 의해서 전신 근육이 긴장되고 저절로 떨 게 됨으로써 체온을 높이려 하고 더울 때는 저절로 근육긴장이 풀려 맥이 풀리고 체온이 떨어지게 된다.
세균에 감염되었을 때 열이 나는 것은 무슨 이유인가?세균이 가지고 있는 내독소 (內毒素) 에 의해서 백혈 수가 자극 받아서 발열물질을 유리하게 되며 이 발열물질이 시상하부의 체온 조절 중추에 작용해서 기준 온도를 올리기 때문이다. 즉 이 발열물질이 체온 기준온도를 39~40°C 로 올리기 위해 근육이 긴장되고 떨게 되며 오한을 느끼게 된다. 그 결과 체온이 오르게 되는 것이다. 이때 해열제를 투여하면 다시 기준 체온이 정상으로 환원되며 체온을 내리기 위해 땀을 흘려서 열을 발산하게 된다. 만일 균 감염이 계속되어 기준체온이 높게 유지된다면 전신근육의 지속적 경련으로 힘이 쇠잔하게 될 뿐만 아니라 체온조절 중추를 포함한 뇌신경세포도 손상을 받아서 기억을 포함한 고위 정신기능의 장애가 동반된다. 흔히 열병을 앓고 난 사람에세 기억감퇴와 고위 정신기능의 쇠퇴를 자주 보게 된다.
이와 같이 우리 뇌 신경세포는 높은 체온에 예민하게 반응을 하여 손상을 입게 된다. 즉 낮은 온도보다는 높은 온도가 뇌신경세포에 더 좋지 않은 영향을 미친다.
미각과 후각을 화학감각이라 한다. 체내에는 화학물질의 농도 변화를 감지하는 많은 화학감수기 (感受器) 가 있다. 일상 생활에서 미각과 후각은 맛나는 것, 향기가 좋은 것 등 향락과 연관 지워 생각하기 쉽지만 원래 후각과 미각은 동물의 생존과 결부된 본질적으로 중요한 감각으로서 먹이의 탐색과 검색을 위해 필요하고, 방어 활동과 성 행동과도 관련이 깊다. 이들 화학감수기는 단백질 수용체로 되어 있으며 화학물질과 반응하여 미각과 후각을 일으킨다.
미각과 후각 감수기 세포는 감각하는 화학물질의 종류가 다르고, 감수성도 다르다. 미각과 후각이 여타 감각과 다른 점은 고도의 적응 능력을 가지는 점과 자극의 질을 식별할 능력, 즉 특정 자극에 대한 감수성이 고도로 분화되어 있는 점이다. 그리고 양적으로 분석할 수 있는 자극강도 범위가 좁아 예민하게 구별 못하는 경우도 많았다. 미각의 자극을 받아들이는 감각기를 맛봉오리 (taste bud) 라 하며 약 2000개 정도 있다. 미각은 네 가지 기본감각, 즉 단 맛 (sweet), 짠 맛 (salty), 신 맛 (acidic), 쓴 맛 (bitter) 등으로 구분한다.
맛봉오리의 흥분은 안면신경과 설인 (舌咽) 신경에 의해 뇌 중추인 대뇌피질의 감각중추에 있는 외측 미각영역으로 전달된다. 이 미각중추가 망가지면 맛을 구별할 수 없다. 우리의 코는 수천 가지 냄새를 식별한다. 콧속에 있는 후각 감수기의 수효는 천만개정도 된다. 여기에서 후각로를 따라 뇌의 이상엽 (梨狀葉), 해마 (海馬) 를 포함한 변연계 (邊緣系) 에 도달되어 최종적으로 냄새를 구별한다. 인간에서는 후각이 다른 동물보다 조금 퇴화되어 있기 때문에 후각기능이 아주 예민하지는 않다.
후각 신호가 변연계에 전달됨으로써 후각의 정서 감정적 요소가 가미된다. 냄새의 질에 따라 즐거움 또는 역겨움 등의 감정이 생긴다. 좋은 향수를 맡으면 강한 무드가 유발되며, 불유쾌한 냄새를 맡으면 재채기 등 방어반사가 일어나고, 암모니아 냄새를 맡으면 반사적으로 호흡이 멈추어진다. 사향향기는 원래 십리 밖의 암사슴을 불러 유혹하는 수사슴 복부 향낭 분비물 냄새이다. 이 향기가 암사슴이 수사슴을 찾게 하고, 암사슴을 성적으로 크게 충동하는 것으로 알려져 있다. 이와 같이 미각과 후각은 생명유지에 기본적인 감각일 뿐만 아니라 보다 나은 맛과 향기를 가진 향료나 음식 개발을 위한 음식문화 창조에 필수적인 감각이다.
평형감각 (平衡感覺) 은 내이 (內耳) 에 있는 두 감각기인 평형반 (平衡班) 과 반고리관에서 느껴지는 감각이다. 인간을 포함한 거의 모든 동물이 생명유전자를 위해 가지고 있는 가장 기본적인 감각중의 하나이다. 가령 개를 들어 뒤집어 떨어뜨려 보면, 떨어지면서 먼저 고개를 돌려 땅을 보며, 땅에 닿기 전 시선을 땅에 고정하고 자세를 바로 잡아 네 발로 사뿐히 땅에 내려선다. 글을 읽으며 고개를 좌우로 기울여 보아도 시선은 글에 고정되어 있다. 이는 모두 평형감각의 장애를 보상하는 반응으로 내이 감각기로부터 시작하는 반응이다. 내이 미로 (迷路) 에는 평형반과 타원낭, 반고리관이 위치하고 있으며 이들은 청신경과 연결되어있다. 이들은 정지된 상태에서 지구인력에 대한 머리와 몸통의 위치 즉 평형 감각을 담당하고 있다. 이들 기관에서의 자극이 최고위 중추인 대뇌피질의 중앙고랑 바로 뒤쪽에 있는 감각중추에 이르러 이 부위에서 이들 기관에서 온 정보를 분석하여 평형과 방위를 인식하며 운동신경에 명령을 내려 자세를 유지하게 한다.
회전운동 감각의 착각으로 정지해 있는 몸이 마치 돌고 있는 것처럼 느끼거나, 외계가 돌아가는 것처럼 느끼는 경우를 현기증 (眩氣症) 이라 한다. 이때 몸의 평형을 유지할 수 없어 넘어진다. 자동차 멀미, 비행기 멀미는 직진운동 감각의 착각에 의하여 일어난다. 미로장해 때에는 안진, 현기증 외에 구토, 발한 등 자율신경계의 증상도 볼 수 있다.
이와 같은 평형감각을 유지하는 기관과 뇌가 없다면 인간은 땅 위에서 두발로 똑바로 걸을 수가 없을 것이며 두 손으로 오늘날과 같은 문명창조는 불가능하였을 것이다.
측두엽 상부에 있는 부위는 감각, 시각과 청각을 연합하고 통합하는 부위이다. 이 영역은 여러 감각을 종합적으로 해석하는 부위이기 때문에 해석영역, 인식영역이라 부르고 있으며 베르니케 (Wernicke) 영역이라고도 부르고 있다. 오른손잡이에서는 왼쪽 뇌에 발달해 있으며 고도의 뇌 기능을 수행한다. 열 사람 중 아홉 사람은 왼쪽 뇌가 우성 뇌이며 약 50%의 신생아는 태어날 때부터 왼쪽 베르니케 영역이 더 크다. 왜 왼쪽 뇌가 우성 뇌이며 오른손잡이가 많은가를 이를 통해 알 수 있다. 어떤 원인으로 우성반구에 있는 베르니케 영역이 손상되면 반대쪽 뇌가 우성반구로 발달된다. 자랄 때부터 크고, 빈번히 사용하게 되므로 이 영역으로 신경전도파가 집중되어 더욱 발달하게 된다. 학습과 기억도 우성 뇌 부위에서 시작하므로 반대쪽보다 일찍 발달하는 것 같다. 인위적으로 자꾸 반대쪽을 사용하게 되면 반대쪽 뇌도 동시에 발달시킬 수 있다. 즉 어느 정도 오른쪽 뇌도 왼쪽 뇌처럼 발달시킬 수가 있다.
또 왼쪽 뇌는 문자나 숫자를 이해하고 계산하는 이성적이고 논리적인데 반해 오른쪽 뇌는 음악이나 그림을 감상하고 비언어적이고 감정적이다. 그러나 이러한 차이점보다는 오른쪽 뇌와 왼쪽 뇌는 앞에서 설명한 바와 같이 같은 점이 훨씬 많다. 운동과 감각, 시각 및 청각을 조절하는 뇌 중추가 같이 위치하고 있다는 점에서 같은 점이 훨씬 많다고 하겠다.
이와 같이 우리는 끊임없는 노력과 각고의 인내로 양쪽 뇌 깊숙이 잠재하고 있는 뇌 기능을 수면위로 떠올려 효과적으로 활용할 수 있다. 이를 통해 우리들은 생활의 효율성을 극대화시킬 수 있다.
수백만 년 전부터 모든 생물의 체내에서 작동해온 생체시계는 지금까지는 미지의 영역이었으나 과학자들의 연구로 점차 모습을 드러내고 있다. 즉 낮과 밤의 주기나 계절의 변화에 따라 우리 몸의 생체시계는 대부분 맞춰져 있으며, 수면과 각성상태, 음식을 섭취하는 일, 모발의 성장, 심장의 박동, 철새의 이동, 근로자의 작업능률등이 생체시계에 영향을 받고 있다. 또한 생체리듬 때문에 약을 먹은 시간에 따라 약효가 다르게 나타나기도 하기 때문에 최고의 약효를 나타내게 하기 위해서는 생체리듬시계를 잘 이용하는 것이 필요하다.
이와 같은 생체시계를 지배하는 인체기관은 뇌의 가운데의 앞부분에 있는 시신경교차 상핵인 것으로 밝혀지고 있다. 따라서 시신경교차 상핵이 파괴되면 신체의 리듬도 사라지게 된다. 이와 같이 시신경이 교차하는 뇌 부위는 생체의 리듬을 조절하는 중앙조절 통제기관이다.
하루 24시간 동안 인간의 생체리듬이 晝夜 明暗주기에 맞추어 활동과 휴식 리듬을 되풀이 하고 있다. 우리의 사고와 행동, 감정을 조절하는 많은 신경전달물질은 각각의 특유한 생체리듬을 가지고 있기 때문에 인간의 사고와 행동, 특히 감정이 시간과 계절의 리듬에 따라서 변화한다. 만물이 생동하고 결실을 맺는 봄과 가을에 사람들의 기분이 많이 좌우를 받는다는 사실로부터 생체리듬의 중요성을 느낄 수 있을 것이다. 혈액 속의 각종 호르몬 양도 시간에 따라 변한다. 성장호르몬과 최유호르몬 (프로락틴) 은 수면시간에는 늘어나고 낮에는 줄어드는 경향을 보이지만 스트레스방어 호르몬인 스테로이드나 카테콜아민은 활동하기 직전인 오전 중에 가장 높다.
복잡한 현대생활에서 오는 스트레스의 증가는 여러 가지 중대한 질병들을 초래하고 있다. 과학자들은 이러한 현대의 질병들이 外的 스트레스 때문에 內的 생체리듬이 파괴되어 나타난다고 믿고 있다. 일시적인 생체리듬의 파괴는 장거리 여행 (특히 비행기여행) 을 빠른 시간 내에 계속해서 했을 때 시차에 적응하지 못한 결과로 나타나기도 한다.
야간작업을 하는 사람들, 밤늦게까지 수험공부에 몰두하는 학생들, 정신적 고민에 쌓여 있는 사람들은 수면리듬이 깨어져서 수면시간이 불규칙하게 될 뿐만 아니라 감정의 리듬과 균형까지도 영향을 받아서 정서가 불안하게 되는 경우가 많기 때문에 수면리듬이 회복되도록 각별히 신경을 기울여 주는 것이 좋다. 최근에는 여러 가지 원인으로 교란된 생체리듬을 정상화시켜 주는 바이오리듬 요법을 통하여 질병을 치료하고자 시도들이 많이 이루어지고 있다. 특히 우리 몸의 생체리듬을 총괄조정하고 있는 시신경교차상핵부의 기능조절이 아주 중요하다.
뇌가 어떻게 바깥 세계에 대한 지식을 획득하는가 하는 것은 중요한 문제이다. 문제의 탐구를 뜻하기 때문이다. 뇌가 위대한 것은 시각대상이 각기 다른 조건에 있다 하더라도 그 대상을 정확히 판별할 수 있다는 점에 있다. 가령 조명조건이 다르면 시각 대상의 표면에서 반사되는 빛의 파장이 변하지만, 그럼에도 불구하고 뇌는 그 시각대상의 색을 정확히 판단할 수 있다. 손짓을 해가면서 말하는 사람의 손의 망막상은 순간순간 변하지만 뇌는 일관되게 그 망막상을 손이라고 판단할 수 있다. 거리가 달라지면 시각대상의 망막상도 달라지지만 그래도 뇌는 그 대상의 진짜 크기를 알 수가 있다. 이와 같이 뇌는 쉴새 없이 변화하는 시각정보의 홍수 속에서 보편적인 시각대상의 특징을 추출해내고 있으며 동시에 시각대상의 해석도 행한다. 해석은 시각과 같이 감각과는 뗄래야 뗄 수 없는 관계를 가진다. 눈앞에 있는 것이 무엇이라는 지식을 얻는데는 뇌는 망막에 비친 대상을 단순히 분석할 뿐 아니라 그 분석에 기초해 시각세계를 뇌속에 적극적으로 조립해 내지 않으면 안되기 때문이다. 뇌는 이 목적을 위해 정교한 신경기구를 만들어냈다. 시각과 관련된 뇌의 구조가 너무도 정교하다.
눈은 삼라만상을 망막 (網膜) 에 맺게 하여 앞을 보는 매우 복잡하고 정교한 장기이다. 눈을 카메라와 견줄 수 있으랴마는 눈과 카메라는 광학적으로 유사한 점이 많다. 앞으로는 빛의 통로인 각막 (角膜) 과 수정체 (水晶體) 가 있고, 안구의 외벽을 구성하는 공막 (鞏膜) 이 보이며, 공막 내측에는 형관성 맥락막 (脈絡膜) 과 빛을 감수하는 망막이 있다. 눈에 빛이 들어오면 수정체는 여러층으로 구성된 망막에 빛의 초점을 맞춘다. 빛에 민감한 신경수용체는 들어온 빛이 자극하는 망막의 표층에 위치하지 않고, 가장 깊은 심층에 위치하고 있다. 즉 빛은 투명한 망막의 여러 층들을 통과하여 심부에 있는 광수용체인 간상체 (nod) 와 추상체 (conc) 세포에 도달한다. 여기에서 다시 표층부에 있는 쌍극신경세포로 보내고, 이어 신경절세포로 간 다음 시신경을 통해 뇌로 들어간다. 간상체 세포는 어두운 빛 (밤의 시각) 을 잘 감지하며 (10조 분의 1와트의 빛도 감지; 80㎞ 떨어진 곳의 성냥불 감지능력) 눈동자의 뒤쪽 벽 측면에 많이 분포하고 있으나 밝은 빛 (낮 시각) 을 잘 감지하는 추상체 세포들은 중앙부의 움푹 들어간 와 (窩) 부위에 많이 밀집해 있다. 따라서 밤에 물체는 간상체가 많이 밀집해 있는 눈동자의 측면에서 잘 보인다. 똑바로 쳐다보지 않아도 물체가 잘 보인다. 우리는 눈을 통해서 본 색깔, 모양, 원근감을 시각중추로 알려진 뇌의 뒷부분에 투사한다. 눈에 들어온 빛이 뇌의 관련된 부분으로 보내질 신호로 변화되는 과정은 놀랄 만한 것이다. 눈에서는 수정체가 망막에서 빛의 초점을 맞추고 이 과정에서 영상은 상하가 바뀐다.
망막에 있는 수용체는 영상을 신경자극으로 바꾸고 이 자극은 시신경을 따라 시신경교차에 도달한다. 이때 눈에서 나오는 시신경의 반이 반대편 뇌로 교차되어 다른 경로를 따라 시각중추에 도달한다. 교차의 결과로 우측 반쪽시야에서 보이는 것은 언제나 뇌의 좌측대뇌반구로 가고 좌측 반쪽시야에서 보이는 것은 무엇이나 우측대뇌반구로 간다. 그러나 뇌 안의 다른 연결을 통해서 다른 쪽이 본 것을 인지할 수 있게 되는 것이다. 뇌 신경계는 시각처리를 분업 체계로 하고 있다. 즉 대뇌피질이나 하부신경계는 시각 대상을 형태, 색, 운동 등으로 나누어 전문 처리하는 영역이 존재하고 있다 머리의 뒤쪽 후두엽에 있는 시각피질이 시각을 관할하는 중추이며 이 부위는 기능에 따라 5개의 영역으로 나누고 있다. 5부위는 각각 형태, 색깔, 운동성분을 구별하는 중추부위이다. 후두엽 바깥쪽 부위는 운동성분을 취급하는 부위로서 여기가 손상을 입으면 대상이 정지해 있을 때는 볼 수 있으나 움직일 때는 볼 수 없다.후두엽 안쪽은 색과 형태를 구분하는 부분으로 여기에 손상을 입으면 색맹이 된다. 그러면 우리는 어떻게 색깔을 느끼는가? 물체란 자기 스스로 어떤 색깔을 가진 것이 아니며 빛을 방출하는 것도 아니다. 뇌가 없다면 세상은 무색일 것이다. 우리가 색깔이라고 느끼는 것은 사실은 빛의 다른 파장들이 우리 뇌의 시각계의 어떤 부분을 자극한 결과다.
추상체라 불리는 광수용체들은 눈의 망막에 있으며 색각에 주로 관여한다. 추상체에는 3가지 유형이 있고 이들은 각각 함유된 화학적 색소의 능력에 따라 광파장의 특정 범위에 민감하다. 추상체의 한 유형은 파란색을 나타내는 어떤 파장을 인지하고 다른 것들은 각기 붉은 색과 녹색을 인식한다. 이들 3가지 색깔은 눈의 기본 색조며 눈의 색조판에서 서로 섞여 모든 다른 색상을 만들어 낸다.
추상체에서 생긴 색 신호는 시각계로 보내지고 여기에 있는 시각경로의 신경세포들이 이들을 배합시킴으로써 우리는 이 세상을 구성하는 약 500만가지 색상의 전체 스펙트럼을 볼 수 있는 것이다. 이들 영역들의 신경세포들은 서로 결합하여 대규모 네트워크를 형성하고 있다. 이 네트워크는 각기 따로 처리된 화상신호를 통합하고 외부의 지시를 얻는데 관여하고 있다. 전에는 뇌에는 '보는' 처리와 '이해하는' 처리를 담당하는 대뇌피질이 다른 위치에 있다고 믿었으나 지금은 복잡하게 상호 연결되어 있음이 밝혀지고 있다.
언어는 인간만이 가지고 있는 고도의 뇌 기능이며 언어가 없었다면 인류 문명의 창조와 발달 특히, 학문의 발달은 불가능하였을 것이다.[언어가 없다면 사고도 없다] 는 말이 있다. 학문을 뜻하는 로고스 (logos) 라는 말은 희랍어에서 유래하였는데 이 말은 이성이라는 뜻과 언어라는 뜻을 함께 가지고 있다. 즉 언어와 사고 나아가서 사람만이 가지고 있는 고유의 기능인 이성은 같은 의미로 해석할 수 있기 때문에 언어의 중요성은 말할 나위 없이 크다고 할 수 있다. 언어로 읽고 들음으로써 지식을 습득하고, 말하거나 씀으로써 전달하고 기록하여 오늘날과 같은 인류문명의 창조와 발달이 가능하였다고 할 수 있다.
뇌의 청각중추는 측두엽 안에 있다. 여기에서 뇌는 소리를 듣고, 크고, 부드럽고, 높고, 낮은 정도에 따라 이 소리를 등록시켜 놓는다. 뇌가 듣는 소리의 세기는 귀에서 포착한 소리 파장의 강도와 연관되고 소리의 고저는 파장의 주파수와 관련된다. 뇌의청각중추는 우리가 받아들인 수많은 소리신호들을 해석하고 다른 것과 비교해 이 소리신호들을 일정한 유형으로 나누며 이러한 과정이 언어 판독에서 가장 중요한 부분이다. 그러나 이들이 고등 뇌중추에 보내지기 전에 두 귀에 들어온 소리신호들은 청신경을 통해 척수의 꼭대기에 있는 뇌간을 통과하게 된다. 거기서 그들은 연속되는 중계 기관을 통과한다. 그 후 많은 신호들은 반대편 뇌에 교차되어 올라가 시상에 모이고 여기서 청각중추로 보내진다. 이 과정의 여러 단계에서 신경세포들은 주어진 신호를 통과시킬 것인지 아닌지를 결정하게 된다. 이 과정이 많은 혼란스런 잡음들을 없앤다.
왼쪽 뇌가 언어를 담당하고 있다. 교통사고나 뇌졸중 등으로 왼쪽 뇌가 망가지면 언어장애 즉 실어증 (失語症, Aphasia) 이 나타난다. 실어증에는 브로카 실어증 (운동성 실어증) 과 베르니케 실어증 (감각성 실어증) 의 두 종류가 있다. 1861년 프랑스의 학자 브로카 (Broca) 는 왼쪽 전두엽의 뒤쪽에 장애를 가진 환자에서 발음능력이 상실된 운동성 실어증이 오는 것을 처음으로 관찰하였기 때문에 언어의 산출과 직접 관련이 있는 전두엽 밑 부분을 브로카부위라고 부르고 있다. 그 후 독일의 학자 베르니케 (Wernicke) 는 브로카가 발견한 운동성 실어증과 다른 실어증인 감각성 실어증을 보고하였다. 이 실어증 환자는 브로카 박사가 기술한 뇌 부위보다 뒤쪽에 있는 측두엽의 장애 때문에 말을 듣고도 그 뜻을 해독하지 못하였다. 따라서 이 부위를 베르니케 부위라고 부른다. 좌측 청각중추에 도달한 신호는 베르니케 영역에 도달하기 전까지는 단지 소음에 불과하다. 이 베르니케 부위가 손상된 사람은 말을 할 수는 있어도 그 뜻을 이해하지 못한다. 브로카의 운동성 실어증은 후두와 입술, 입, 호흡계, 기타 목소리를 내는데 필요한 발성근을 조절하는 뇌 부위가 손상되었기 때문에 듣거나 읽고서 언어를 해독하기는 하지만 말하거나 쓰기와 같은 언어의 표현이 불가능하다. 단어를 찾아낼 수는 있지만 말을 조직적으로 하기 위한 능력이 소실되어 있다. 반면 베르니케의 감각성 실어증은 측두엽 위쪽의 장애로 일어나며 말과 글을 듣고 볼 뿐 그 뜻을 분석하는 능력이 소실된다. 생각은 이루어지지만 그 생각을 표현하기 위해 적당한 낱말들로 이어가기가 힘들어 진다.
이와 같이 우리 뇌의 어느 부분도 중요하지 않은 부분이 없지만 특히 언어와 관계된 브로카 영역과 베르니케 영역은 언어와 문화창조의 핵심부라 할 수 있다.
신경세포는 그들이 가지고 있는 세포질 돌기의 수에 따라서 나눈다. 가장 간단한 경우는 세포질이 축삭 (axon) 이라는 한 개의 돌기만을 가지는 단극 (單極) 신경세포 (nuipolar cell) 이다. 좋은 예는 비척수동물의 신경세포와 척추동물의 등뼈 속의 척수에 있는 배근 신경절 (背根神經節; dorsal root ganglia) 에 존재하는 감각 신경세포이다. 두 번째는 두 개의 돌기 (축삭과 수상돌기) 를 가지고 있는 세포, 즉 양극(兩極) 신경세포 (bipolar neuron) 이다. 망막과 후각 점막에 있는 감각수용체 신경세포와 청각 신경세포, 조그마한 과립세포가 여기에 속한다. 세 번째는 한 개의 축삭돌기와 한 개 이상의 수상돌기를 가지는 다극 (多極) 신경세포 (multipolar cell) 이다. 대부분의 척추동물의 신경세포는 여기에 속한다. 신경세포 축삭돌기의 형태를 유지하는 데는 부수 요소인 신경교세포 (神經膠細胞) 가 깊이 관여하고 있다. 이 세포의 이름은 그리스어의 아교라는 말의 glue 에서 膠 를 따왔다. 신경교세포에는 성상세포 (星狀세포; astrocyte) 와 올리고덴드로사이트 (oligodendrocyte) 가 속한 대 (大) 글리아 세포와 외부에서 들어온 이물질을 잡아먹는 소 (小) 글리아 세포가 있다. 이중에서 가장 중요한 두 가지의 대글리아 세포를 보면 첫째는 성상세포 (아스트로사이트; astrocyte) 인데, 이 이름은 광학 현미경하에서 '별' 모양으로 생겼기 때문에 붙여졌다. 이 세포는 축삭돌기와 수상돌기가 있는 부위에 존재하며, 혈관 벽을 싸고 있다. 이 세포의 구조적 특징으로 볼 때 신경 전도면 사이에서 절연 작용을 나타내며, 신경 말단과 수상돌기의 기능적 단위를 둘러싸서 분리시키는 작용을 하는 것 같다.
두 번째 세포 형태는 올리고덴드로사이트 (oligodendrocyte) 이다. 이 세포는 신경 세포체 주위에 있으며, 말초 신경계에서는 스완세포 (Schwann cell) 주위에 붙어 있기 때문에, 이런 부위에서는 위성 세포 (satellite cell) 라 불린다. 올리고덴드로사이트의 세포질은 거친 내형질 망상체 (內形質網狀體; rough endoplasmic reticulum) 를 가지고 있으며, 가장 뚜렷한 특징은 축삭돌기 주위를 세포막으로 동심원 형태로 감싸고 있다는 것이다. 이 동심원 층이 간격이 없이 매우 가깝게 감싸고 있기 때문에 세포질이 완전히 빠져나가, 처음에 싸고 있던 막들이 서로 합쳐져서 완전히 반지 모양의 미엘린 초 (myelin sheath) 를 나타내고 있다. 수 센티미터 (㎝) 길이를 뻗어 나가는 축삭돌기를 따라서 많은 올리고덴드로사이트 세포들이 미엘린 초를 형성하여 유수신경을 형성하고 있다. 축삭돌기를 감싸고 있는 사이사이에는 감싸지 않은 부분이 존재하고 있는데, 이 부위를 랑비에르절 (node of Ranvier) 이라 부른다. 많은 중추 신경계 축삭돌기와 말초 자율 신경계의 어떤 부위는 미엘린 초를 가지고 있지 않다. 이 신경을 무수신경이라 부른다. 축삭돌기와 올리고덴드로사이트 사이의 밀접한 관계로 볼 때 올리고덴드로사이트는 신경 세포의 성장에 기여하고 있는 것 같다. 그러나 신경교세포는 외상이나 수술에 의한 손상으로 신경세포체에서 축삭돌기가 달려 나갈 때 이 축삭돌기를 지지해 주지 못하고 있다. 뇌 손상 후 축삭돌기가 변성되어 나가는 것을 염색하여 볼 수 있는데, 이 염색 방법으로 신경교세포가 축삭돌기를 지지해 주지 못한다는 사실이 발견되었다.
중추신경계에는 글리아 세포가 신경세포보다 10배~50배 이상 많다. 이 글리아 세포는 모양이 독특하여 신경세포와 달리 전기적 시그날을 전파하지 못하다. 신경세포체와 축삭돌기 사이에 위치하고 있으며 다음과 같은 6가지의 기능을 하는 것으로 알려지고 있다.
① 글리아는 아교 (Glue) 라는 뜻의 그리스어에서 유래한 것처럼 뇌의 구조를 지지해주는 세포이다. 즉 다른 조직에서 보는 결체조직 (connective tissue) 의 역할을 한다. 또한 같은 기능을 하는 신경세포들의 집단을 분리해주거나 신경세포를 절연시켜주는 역할도 수행한다.
② 어떤 글리아 세포들은 손상으로 인하여 죽은 신경세포들을 제거해주는 역할도 한다.
③ 어떤 글리아 세포는 축색을 둘러싸서 절연효과를 나타내는 미엘린 (myelin) 을 생산한다.
④ 글리아 세포들은 세포외액의 K+농도의 급격한 변화를 완충시켜주며 흥분에 의해서 신경세포에서 유리된 신경전달 물질을 제거해주는 역할도 한다.
⑤ 성장기에는 어떤 종류의 글리아 세포들은 신경세포의 성장과 이동을 안내하고 도와준다.
⑥ 어떤 글리아 세포들은 신경세포에 영양을 공급해준다.
신경섬유는 신경계의 전기 줄에 해당하는 것이라 볼 수 있다. 이를 따라 메시지를 지닌 신경 흥분이 신체의 한 곳에서 다른 먼 곳으로 전달된다. 그렇다고 하여 신경섬유가 전선과 같이 단순히 전류가 흐르는 전도체는 아니다. 구리로 된 전선을 따라 전류가 흐를 때는 대략 광속에 가까운 속도로 흐르지만 신경섬유를 따라 흥분이 전도되는 속도는 기껏 빨라야 매초 100m에 지나지 않는다. 더욱이 축삭 속의 전기 저항은 구리줄보다 약 1억 배나 크고, 이엘린껍질의 전기적 절연성은 질이 좋은 전선 껍질의 100만 분의 1에 불과하다. 그러므로 만약 신경섬유가 단순히 전기적 도체로만 작용한다면 신경흥분은 단 몇 ㎜ 도 나아가지 못할 것이다. 이렇듯 신경섬유는 전기적 도체로서가 아니라 이것이 갖추고 잇는 증폭계와 릴레이계에 의하여 신경흥분을 효과적으로 전달한다.
신경섬유의 내부와 외부를 비교해 보면 뚜렷한 전기 화학적 차이가 있다. 신경섬유의 내부에는 각종 유기화합물이 많으며, 특히 K+농도가 높다. 신경 섬유 외부는 세포외액에 공통된 Na⁺과 Cl- 농도가 높다. 그리하여 세포안 K+농도는 세포 밖보다 약 30배나 높으며, 세포 밖 Na⁺농도는 세포 안보다 약 10배나 높다. 이러한 농도 차이로 인하여 세포막이 허락한다면K+은 세포 밖으로, Na+은 세포 안으로 확산되려고 한다. 물론 여기서 세포 안 유기화합물은 세포막을 쉽게 투과하지 못한다. 그러나 Na+ 은 투과되기가 매우 어려워 Na+의 투과성은 K+의 약 1/1000에 불과하다. 그러므로 K+은 세포 내외에 이미 설정된 30대 1의 높은 농도 기울기로 말미암아 세포 밖으로 확산되어 나오려고 한다. 전기적으로 양성을 띤 K+이 확산되어 나오려 하는 힘은 곧 세포 속을 전기적 음성으로 만들 게 되는 것이다. 그렇게 되면 이 세포 내 음전기는 K+을 세포 밖으로 나가지 못하게 안에 붙잡아 두는 힘이 되기도 한다. 그리하여 세포 내 음하전으로 K+을 세포 내에 붙잡아 두는 힘과 농도 기울기로 인해 밖으로 확산되어 나가려는 힘이 어느 시점에서 균형을 이루게 된다. 이때 세포 안은 대략 -90㎷의 전위를 띠게 되는데, 이 전위를 가리켜 막전위 (膜電位, membrane potential) 혹은 안정전위 (安靜電位, resting potential) 라 한다. 이때 세포 속과 밖은 전위차가 나타나기 때문에 분극 (分極) 이 되어 있다고 할 수 있다. 막전위는 K+이 세포 밖으로 확산되어 나가려는 힘이 연유된 전위이다. 만약 안정 세포막이 K+대신 Na+에 대한 투과성이 높다면 어떠할까? 농도 기울기는 세포 밖에서 안으로 향하고 있으므로 이번에는 오히려 세포 안이 양하전으로 기울 게 될 것이다.
이와 같이 여러 이온의 세포막을 통한 투과성이 변화함으로써 막전위가 변동할 수 있는데, 이 변동은 신경섬유가 흥분하였을 때 일어나는 변화이기도 하다. 신경섬유를 전기 자극하여 막전위를 -90㎷ 에서 0㎷ 로 끌어올리면 세포 속의 음전위가 없어지기 때문에 세포 속과 밖의 분극이 없어지게 된다. 이것을 탈분극 (脫分極, depolarization) 이라 한다.막전위가 오르는 것과 때를 같이하여 세포막은 K+을 제쳐놓고 이번에는 Na+에 대한 투과성이 갑자기 증가한다. Na+의 농도 기울기는 밖에서 안으로 향해 있으므로 Na+은 세포 내로 몰려든다. 그러면 또 막전위는 더욱 더 오르게 되며 Na+투과성은 한층 더 높아진다. 즉 어느 범위 내에서는 막전위가 올라 갈수록 Na+의 투과성은 더욱 높아져 많은 Na+이 세포 내로 들어가게 된다. 이리하여 마치 세포막이 갑자기 Na+에 대해 문을 활짝 여는 것처럼 Na+이 세포 속으로 몰려 들어가게 되며, 매우 짧은 시간에 -90㎷ 이전 안정전위가 +30㎷ 로 뛰어 오르게 된다. 이것이 활동전위 (活動電位, action potential) 라 부르며 신경 흥분의 첫 단계이다.
활동전위가 발생된 신경섬유 부분은 세포 내 하전이 음성에서 양성으로 바뀌었지만 인접 안정 부분은 전기적 음성이어서 전류는 섬유 속 + 부분에서 -부분으로 흐르게 된다. 이 전류는 세포막을 뚫고 나와 세포외액을 돌아 다시 흥분된 제자리에 되돌아오는 전류의 회로를 만든다. 이 국소 전류는 활동전위가 발생된 바로 옆 안정 부분 막전위에 영향을 미쳐 이곳도 탈분극 시키고 같은 방식으로 바로 옆 다음 부분도 Na+에 대해 문을 열어 주어 활동전위가 유발되게 한다. 즉 한 곳에서 활동전위가 발생되면 그 마디가 다음 마디를 자극하여 흥분시키고, 다시 다음 마디로 활동전위가 릴레이되어 나아가므로 축삭돌기 끝에서 끝으로 흥분이 전달되게 한다. 그러므로 신경의 흥분 전도는 도화선의 연소와 흡사하다고 할 수 있다. Na+에 대한 투과성이 높아져 활동전위가 발생하였으나 Na+투과성은 곧 쇠퇴하고 이번에는 K+에 대한 투과성이 높아져 K+의 문을 활짝 열어 준다. 그러면 K+은 세포 밖으로 확산되어 활동저위는 다시 안정전위로 회복된다. 이 과정을 가리켜 재분극 (再分極, repolarization) 이라 한다. 재분극 된 세포막은 다시 안정시의 투과성을 찾게 된다. 이상 활동전위는 매우 빠른 과정이어서 그 시간은 0.001초에 불과하다.
신경 흥분의 전도, 즉 활동전위가 전파되어 나아감을 가리켜 충격파라 한다. 충격파는 신경이 지니고 있는 본연의 성질에 기인된 것이다. 그러므로 충격파가 나아갈 때 신경섬유에서 일어나는 물리 화학적 변화의 크기와 속도는 자극의 종류나 강도와 관계없이 언제나 일정하다. 우리가 화약을 가볍게 치거나 세게 치거나 일단 화약이 터질 때는 그 폭발력은 언제나 같은 법이며, 화약을 세게 친다 하여 더 빨리 크게 폭발하지는 않는 이치와 같다.
복어의 난소와 간에 함유된 복어독 (tetrodotoxin, TTX) 은 신경섬유가 자극을 받아도 세포막이 Na+에 대한 투과성을 높여 Na+에 대해 문을 활짝 열지 못하게 하는 독약이다. 이른 바 Na+의 통로를 막아 버리는 약물이다. 그리하여 신경섬유에 활동전위가 발생되지 않게 하며, 신경 흥분이 전도되지 않게 된다. 이렇게 충격파가 발생되지 않게 되면 근육이 마비되고, 호흡근 마비로 호흡이 멎어 죽게 된다. 결국 신경의 흥분성과 전도성은 오직 신경막의 특유한 투과정 그리고 세포 밖의 높은 Na+농도와 세포 속의 높은 K+농도에 유래된 것이다. 여기에 포도당, ATP는 에너지원으로서 Na-K펌프를 가동시켜 세포 내외의 특유한 Na+ , K+의 농도 분포를 유지하게 된다.
무게가 평균 1300g이며 몸무게의 2%, 펼치면 2300㎠로 신문지 한 장 정도의 적은 표면적을 가지고 있지만, 우리 몸의 산소 소모량과 혈류량은 10배인 20%를 차지하고 있는 이 중요한 구조가 인간 존재의 본질을 표현하고 있다. 뇌의 극히 작은 부분, 단 몇 밀리미터 정도라도 손상된다면 인생이 영원히 바뀌어 버릴 수도 있으며 뇌의 기능이 심하게 손상된 인간은 식물과 같이 사고와 운동을 할 수 없기 때문에 식물 인간으로 불려지며 인간 생명 존재 가치가 근본적으로 의심을 받게 된다. 이렇게 놀라운 해부학적 존재를 이루고 있는 뇌에 대하여 인류 역사가 시작된 이래로 인간은 뇌의 신비를 밝히기 위해서 끊임없는 노력을 지속하여 왔다. 우리가 아무리 복잡한 정보체계를 상상한다 해도, 수백 억~수천 억개에 이르는 무수한 신경세포가 거미줄처럼 서로 다른 수천 개의 신경세포와 연결되어 교신을 하고 있는 뇌의 복잡성에는 따라가지 못 할 것이다.
한 개의 신경세포는 수천 개의 신경세포와 교신을 주고받고 있는데 이러한 교신을 담당하고 있는 본체가 바로 화학물질인 신경전달물질 (neurotransmitter) 이라는 사실이 밝혀진 것이 금세기에 이루어진 가장 획기적인 발견중의 하나이다. 20세기 초까지만 하더라도 신경세포와 신경세포 사이에는 세포질이 서로 전기 줄처럼 연결되어 흥분이 전도되는 것으로 생각하였다. 그러나 현미경으로 자세히 관찰한 결과 신경세포 사이에는 항상 일정한 간격 (틈) 이 존재한다는 사실이 밝혀졌으며, 이러한 간격을 뛰어 넘어서 흥분이 전달되기 위해서는 어떤 매개물질 (메신저) 의 존재가 필요하다는 자연적인 추론이 나오게 되었다.
1921년 오토뢰비 박사는 미주 신경 (vagus nerve) 이 붙어 있는 개구리 심장과 미주신경을 제거한 개구리 심장을 준비하여 각각 링거액에 담그고 서로 링거액을 통하게 연결시켰다. 첫 번째 심장에 붙어있는 미주신경을 자극하였을 때 심장의 박동이 느려졌으며 미주신경이 없는 두 번째 심장의 박동도 느려졌다. 즉 첫 번째 심장에 붙어있는 미주신경을 자극하였을 때 이 신경의 말단에서 어떤 화학물질이 유리되어져 나와 링거액을 통해 신경이 없는 두 번째 심장에 직접 영향을 미친다는 사실을 밝힘으로써 신경전달물질의 존재를 처음으로 증명하였으며 이 신경전달물질을 미주신경물질 (vagusstuff) 로 명명하였다. 이후 이 물질은 아세틸콜린임이 밝혀졌으며 현재까지 뇌에는 40여 종류가 넘는 신경전달물질이 발견되고 있다.
우리는 흔히 이 세계를 눈에 보이는 물질세계와 보이지 않는 정신세계로 나누고 있다. 정신세계를 움직이고 조절하는 것도 물질로 이루어진 화학적 신경전달물질에 의해서 이루어지고 있다는 사실은 많은 것을 시사해 주고 있다. 현재 복잡한 정신세계, 마음의 세계를 눈에 보이는 과학적 개념으로 모두를 설명할 수는 없지만 과학이 발달해 감에 따라 현재 보이지 않는 세계, 추상적인 세계도 구체적인 현실세계로 볼 수 있을 것이다. 눈에 보이는 세계와 보이지 않는 세계의 정의가 과학적으로 상당히 애매해 지고 있는 것이 사실이다. 과거에 존재를 볼 수 없었던 많은 것들이 현재 그 존재를 인정 받고 있다.
정신분열병, 우울병, 신경증, 파킨슨병, 무도병, 간질, 자폐증, 수면장애 등과 같은 중요한 신경정신계 질환이 특정 신경전달물질계의 기능과다 및 기능감소로 생긴다는 사실이 밝혀지고 있다.
뿐만 아니라 현재 크나큰 사회적 문제를 야기하고 있는 히로뽕, 코카인, LSD 같은 환각제도 신경전달물질계에 영향을 미침으로써 환각 및 행동의 이상을 보인다고 생각하고 있다. 즉, 뇌의 모든 기능은 많은 종류의 신경전달물질에 의해서 이루어진다. 다시 말해서 인간을 인간답게 만들며, 인간활동의 최고 주체이며, 인류문화 창조의 근원을 형성케 했던 중심이 신경전달물질이라고 말해도 과언이 아니다. 중요한 역사적 사건의 주체들, 인류에 큰 타격을 주었던 전쟁을 일으켰던 사람들의 생각과 사상, 행동을 이해하기 위해서는 이들의 신경전달물질계가 어떻게 다른가 하는 연구가 필수적일 것이다. 고도의 정신기능, 감정, 운동 및 감각기능을 하기 위하여 얼마나 만은 신경전달 물질이 뇌에 존재하는지 아무도 모르고 있다. 앞으로 과학이 발달함에 따라 서로 다른 기능을 하고 있는 많은 신경전달물질들이 끊임없이 발견될 것이며 이 신경전달물질계의 특성을 밝힘으로써 인간정체의 본질이 규명될 수 있을 것이다.
[표 1] 신경전달물질
|
농도 |
작용 |
|
1-15u moles/g조직 |
이온향성(이온向性) Type Ⅰ 글루탐산 (Glutamate) (이온의 이동을 통해서 신경 흥분을 전도한다) Ca2+,Na+,K+ (아미노산) 아스파르트산 (Aspartate) Cl- GABA Cl- 글리신 (Glycine) 빠른 작용 세포 : 뇌 여러 부위 |
|
농도 |
작용 |
|
1-25n moles/g |
대사향성 (代謝向性 : G단백을 Type Ⅱ 아세틸콜린 통해 작용을 나타냄) 느린작용 (아민) 도파민 이차전달자와 연결 노르에피네프린 세포체 : 뇌저부위 에피네프린 5-HT(세로로닌) 히스타민 |
|
농도 |
작용 |
|
1-500n moles/g |
대사향성 Type Ⅲ CCK(470) Type Ⅰ과 Ⅱ와 공존(펩티드) 엔케팔린 metenk(350) 하고 있는 공존전달물질(共存 傳達物質) (신경조절물질) 세포체: 시상하부 여러 뇌 부위 P물질(SP)(100) VIP(40) 소마토스타틴(SOM) LHRH(7) 디놀핀(Dynorphin)(4) 바소프레신 (Vasopresin)(3) β-endolpine(엔돌핀) ACTH α-MSH(멜라닌세포자극호르몬) TRH(갑상선자극유리호르몬) |
인간의 모든 정신작용과 갖가지 활동은 신경전달물질이라고 하는 화학물질에 의해서 이루어지고 있다. 말초조직에 작용하고 있는 많은 호르몬도 뇌신경세포에 존재하며 이 호르몬도 뇌에서는 신경전달물질로서 작용하고 있다. 신경전달 물질에 의해 정보전달이 이루어지고 있는 신경세포와 신경세포의 접합부위를 '시냅스' 라고 부르며 이 시냅스는 전깃줄처럼 이어져 있는 것이 아니라 일정한 간격을 가지고 있다. 즉 이 시냅스 간격을 뛰어넘어 정보를 전달해 주는 메신저 물질이 신경전달물질인 것이다.
신경전달물질 (호르몬 포함) 은 신경섬유 말단부의 조그마한 주머니인 소포체에 저장되어 있다가 신경정보가 전기적 신호로 신경섬유막을 통해 말단부로 전파되어 오면 이 조그마한 주머니가 신경세포막과 접합한 후 터져서 신경전달물질이 시냅스 간격에 유리된다. 유리된 전달물질은 2만 분의 1밀리미터 정도 되는 짧은 간격을 흘러서 다음 신경세포막에 도달되며 세포막에 있는 특수한 구조와 결합함으로써 흥분이 전달된다. 이 특수한 구조는 정보를 받아들이는 물질이라는 의미에서 受用體 (Receptor) 라고 부르며 단백질로 구성되어 있다. 즉 신경전달물질은 일종의 열쇠이며 이를 받아들이는 수용체는 열쇠 구멍에 해당되기 대문에 전달물질이라고 하는 열쇠가 수용체라고 하는 열쇠구멍에 맞게 결합함으로써 다음 신경세포막에 있는 대문이 열려 정보가 전달될 수 있는 것이다. 신경전달물질은 각자 특유의 수용체분자하고만 결합하여 특정 정보 전달을 한다. 즉, 신경정보를 가지고 있는 신경전달물질이라고 하는 화학분자와 그 정보를 받아들이는 수용체라고 하는 특수 탄백질 분자의 상호 결합으로 고도의 정신기능에서부터 행동, 감정에 이르기까지 모든 것이 결정되는 것이다. 유리된 신경전달물질이 신경세포막에 있는 수용체 단백질과 결합하면 수용체 분자 옆에 있는 이온통로 (이온채널) 가 활성화되어 Na+, Ca++ 과 같은 양이온 염소이온 (Cl-) 과 같은 음이온의 세포내로의 이동이 증가된다. 평상시 세포 내는 -60㎷에서 -90㎷의 음전하를 띠고 있으나 양이온 이동의 증가로 양전하를 띠게 되면 신경세포는 흥분 발사를 하게된다. 이때 연소이온과 같은 음이온이 세포 내로 들어오게 되면 세포 내의 음전하는 더욱 커지게 되어 신경세포의 흥분은 억제되는 것이다. 신경세포를 흥분시키는 전달물질로는 글루탐산 (glutamic acid) 이, 억제시키는 전달물질로는 GABA (r-aminobutyric acid) 가 대표적이다.
신경전달물질이 적절히 유리된다고 하더라도 이와 결합하는 수용체가 적절한 기능을 하지 못한다면 신경정보는 효율적으로 전달이 되지 못한다. 이런 의미에서 전달물질과 수용체는 뇌에서 활동하는 男女주역인 것이다. 어떤 이유로 전달 물질이 유리가 적어지면 수용체 수는 증가되며 반대로 유리가 너무 많아지면 수용체 수는 줄어들어 우리 뇌의 기능이 일정하게 恒常性 을 유지하게되는 것이다. 이러한 항성성이 깨어질 때 여러 가지 신경정신질환이 발생된다. 대표적인 예를 들면 도파민 신경전달물질이 유리되어져 나오는 신경세포가 망가지면, 수용체는 정상이나 수용체와 결합하는 도파민 전달물질이 없기 때문에 도파민이 매개하는 운동기능이 상실됨으로써 '파킨슨병' 이 생기고 반대로 도파민 수용체가 과도한 활동을 하면 '정신분열병' 이 생기는 것으로 생각하고 있다. 따라서 파킨슨병을 치료하는 방법은 도파민 신경전달물질의 합성을 증가시키는 약제를 사용하는 것이며, 정신분열병을 치료하는 방법은 도파민 수용체의 기능을 차단하는 약물을 사용하는 것이다. 앞으로 뇌의 두 주역인 신경전달물질과 수용체의 특성에 관한 연구가 과학의 첨단연구가 될 것이며, 이 두 주역의 정체 해명으로 인간정신의 해명, 나아가 생명의 수수께끼를 풀 수 있는 거보를 내딛게 될 것이다.
매일 일어나고 있는 거의 모든 현실은 우리와는 별개로 일어나고 있지만 우리가 그 실체를 제대로 인식하고 파악할 때에만 참모습으로 우리에게 다가온다. 이러한 현실세계의 실체를 인식하고 파악하는 것은 전적으로 우리 뇌의 기능에 속하고 있다. 건강하게 우리 뇌가 살아서 작동하는 한 우리는 이 우주의 모든 것을 제대로 인식하고 탐구할 수 있다. 그러나 어떤 사람의 뇌가 실제 세계에 존재하고 있는 것으로 받아들이지 않으면 비록 존재하고 있는 것도 그 사람에게는 존재하고 있지 않는 것으로 된다. 또한 어떤 사람의 뇌가 실제 세계에 존재하고 있지 않는 것을 실제 존재하고 있는 것으로 받아들일 때는 그 사람에게는 존재의의가 있게 된다. 사고나 전쟁에서 한쪽 다리를 잃은 사람들이 마치 다리가 다시 붙어있는 것과 같은 감각을 느끼거나 다리에 벌레가 기어가는 것과 같은 간지러움도 경험한다. 이와 반대로 뇌졸중과 같은 질병을 앓고 있는 사람이 한쪽 다리가 멀쩡하게 붙어있는데도 없다고 느끼는 경우도 있다. 이와 같이 내가 생생한 현실을 체험하고 못하고 하는 것은 나의 뇌 속에서 일어나는 현상에 전적으로 의존하는 것이지 반드시 실제 세계 속의 존재여부와는 관계없다. 그래서 삶의 모든 현상들은 순간 순간 뇌에서 일어나고 있는 생물학적 과정에 불과하며 인간의 정신과 육체는 독립적인 실체가 아니라 하나라고 하는 一元論 (Monism) 또는 唯物論 (Materialism) 이 정신과 육체가 서로 독립되어 다른 존재 양식을 가지고 있다고 하는 二元論 (Dualism) 보다 설득력 있게 받아들여지고 있다. 뇌사나 식물인간의 경우에서 보는 것처럼 뇌가 죽으면 인간으로서의 존재 가치가 상실되고 어떤 정신기능이나 마음이 창출될 수 없기 때문에 뇌는 生 과 死, 인간과 다른 생물을 구분 짓는 유일한 가치기준이 되고 있으며, 우리의 정신과 마음, 행동, 성격이 뇌로부터 나온다는 점은 모두 인정하고 있다.
뇌신경세포를 다른 사람의 뇌에 이식하여 주면 그 사람의 정신과 마음이 반영될 수 있는가 없는가 하는 문제, 즉 뇌신경세포를 기증한 사람의 정신과 마음이 뇌세포를 받은 사람의 정신과 마음에 스며들어서 서로 싸우게 되지는 않을까 하는 문제는 간단하지는 않다. 그러나 뇌가 그 사람의 특성을 결정해주는 유일한 장기이기 때문에 이식한 뇌의 특성이 그 사람에게도 상당부분 나타날 수 있을 것이다. 로보캅이라는 영화에서 이식한 경관의 뇌가 옛날의 일을 회상할 수 있고 경관의 감정을 어느 정도 표출할 수 있었던 것도 이런 이유 때문이다. 즉 뇌가 곧 나이며 내가 곧 뇌인 것이다 (I am the brain).
정신과 마음은 무엇이며 어떻게 다른가?
정신은 집단의 생명을 유지시켜주는 궁극의 원리로서 보다 집단적이고 객관적인 반면 마음은 생명체 개개가 가지고 있는 정서적이고 감정적인 측면에서의 차원 높은 원리로서 보다 정서적이고 감정적이며 개인적인 면을 말한다. 정신을 고귀하고 마음은 따뜻하고 부드러우며 아름답다고 보통 이야기 하나 반대로 정신이 따뜻하고 부드럽다고 표현하지 않는다. 정신은 창조와 고등정신기능을 관찰하는 대뇌 신피질부의 시경세포 활동으로 나오나 마음은 이 신피질부의 활동과 감정을 조절하는 고피질부의 활동이 첨가되어 나온다고 볼 수 있다.
최근 인식 (認識), 인지 (認知) 를 연구하는 인지과학 (Cognitive Science) 이 새로 태동되어 신경계를 연구하는 신경과학 (Neuroscience) 과 같이 발전하고 있다. 신경과학은 인간의 정신과 마음을 구성하는 하드웨어인 뇌의 구조와 소프트웨어인 화학, 생물학 및 행동 특성을 연구하는 반면 인지과학은 인간의 정신과 마음을 구성하는 기능적 측면 즉 소프트웨어 쪽만을 주로 연구하는 학문이다. 매일 보고 듣고 느끼는 모든 과정들을 뇌 신경세포는 특유한 방식으로 수집하여 중추로 전달하고 중추에서는 이를 신경세포 회로망 속에 새겨 넣어 적절한 변형과정을 거친 다음 표상 (representation) 으로 나타낸다. 여기에 병렬연결 방식을 가진 신경 컴퓨터를 개발하여 한 단계 위에서 검토하고 판단함으로서 인간과 같은 자성능력을 가진 인공지능 (Artificial Intelligence) 경컴퓨터, 인공지능 분야는 빠른 속도로 발전할 것이며 나아가서 인간을 닮은 인조인간을 어느 정도는 제조할 수 있을 것이다. 인간 뇌의 기능을 가진 인조인간의 개발은 인간의 정체성 (Identity) 및 윤리에 크나큰 변혁을 초래하게 될 것이다. 뇌에 관한 인간의 지식이 깊어질수록 인문사회과학분야, 자연과학분야 나아가서 산업계에는 혁명이 일어 날 것이다.
인간의 합리적 사고와 이성에 몰두해있던 프란시스 베이컨의 시대에는 인간의 감정은 도외시되었다. 문제를 인지하고 합리적인 판단을 통해 문제를 해결하는 인간이성의 논리성을 규명하는 것이 곧 인간적 본질을 파악하는 중심적 과제가 되어왔었다. 따라서 감정에서 파생된 기쁨과 슬픔 공포와 불안 혐오감 같은 것은 과학자들의 연구영역에서 사소하고 별 것 아닌 것들로 여겨져 왔다. 그러나 사람들의 동기를 유발시키고 구체적인 행동 및 삶의 양식들을 꾸려 나가는 주요 원천이 합리적 이성에만 있지 않다는 사실이 밝혀지면서 인간의 감정이 중심적인 연구대상으로 자리잡아가고 있다. 즉 감정은 인간정신의 사소한 파생물이 아니고 기억과 판단, 학습 등 고도의 이성적 사고와도 관련지어 있다는 것이다.
이성과 더불어 인간본질을 구성하는 양 축의 하나로 새롭게 조명되고 있는 감정에 대한 연구는 감정이란 정신현상이 인체의 어느 부분과 관련되어 있는지에 주로 초점이 모아지고 있다 20세기에 들어와서도 대부분 인간의 사고과정에 주목해왔던 뇌를 연구하는 과학자들은 감정의 생물학적 근거도 뇌의 특정부위를 연구함으로써 감정의 진원지를 알 수 있는 것으로 보고 있다. 감정현상과 뇌 영역의 관련성에 대한 연구결과 대뇌피질 안쪽에 오래된 뇌인 변연계라는 조직이 감정과 관련된 정보를 받아들이고 신체적 반응까지도 조절한다는 사실이 밝혀졌다. 인간의 생체조직과 감정과의 연관성에 대한 논의에서 대표적인 것은 뇌의 좌우 반구가 각기 다른 감정적 반응을 담당하고 있다는 것이다. 예를 들면 왼쪽 뇌가 손상된 경우는 웃는 얼굴표정을 짓지 못한다고 한다. 보다 구체적으로 공포나 혐오와 같은 부정적인 감정표현이 요구될 때 오른쪽 뇌의 활동이 활발해지고 유쾌하고 행복한 기분에 젖어들 때는 왼쪽 뇌의 작용이 왕성해 진다는 것이다.
한편 손톱 크기 만한 편도핵 (扁桃核) 이 뇌의 각부분과 작용하면서 일종의 감정을 주관하는 센터가 아닐까하는 연구도 있다. 이미 지난 30년대에 원숭이를 대상을 연구한 내용에 따르면 편도핵이 손상되면 외부의 자극에 대한 반응을 망각하게 돼 위험스런 상황인지를 전혀 알지 못하고 통상 놀랄 만한 상황에서도 전혀 감정표현을 하지 못하는 경우가 발견되었다. 편도핵이 감정적 판단을 하는데 일차적으로 작용하는 신체요소라고 지적되고 있는데 일례로 숲 속을 걸으면서 무슨 소리를 들었을 때 총소리로 생각해서 불안과 공포를 느낄 것인지, 나무 부러지는 소리이기 때문에 아무 동요를 느끼지 않을 것인지에 대해 편도핵이 먼저 반응을 한다. 이러한 논의는 감각 정보가 뇌로 먼저 전달된 후 하부 신경계통으로 반응명령을 내린다는 과거의 과학자들 생각과는 다른 내용이다. 실제로 대뇌피질 등이 파괴돼도 불안 공포와 같은 원초적 감정표현은 편도핵을 통한 긴급회로에 의해서도 이루어질 수 있다는 것이다. 이런 논의는 감정처리가 논리적 판단 이전에 무의식적으로 일어난다는 견해에 대한 생물학적 근거를 제시한다. 즉 감정은 사고보다 보다 원초적인 기능으로 고등동물은 정도의 차이는 있지만 공통으로 고차원적인 감정은 고도의 사고기능과 밀접하게 연결되어 있지만 본능적인 공포나 놀람과 같은 원시적인 감정은 대뇌피질까지 연결되지 않고 하부 뇌에서 반사적으로 이루어진다. 명랑하면 우울할 때보다 두뇌능력이 우수해진다. 어떤 문제를 풀기 위해 감각을 기록하고 이를 처리 활용하는 인간의 기억작용이 얼마나 우수하게 발휘되는가는 기분에 따라 크게 좌우되기 때문이다. 특히 복잡한 과제의 해결에 있어서는 명랑한 사람이 우울한 사람보다 훨씬 탁월한 능력을 보이게 된다. 이 같은 사실은 최근 독일 괴팅겐대학 심리학과 게르트 위에 교수의 실험연구를 통해서 확인됐다. 실험의 핵심은 대상을 기분상태에 따라 명랑한 그룹과 우울한 그룹으로 나누어 자연과학학술도서를 읽게 하였다. 책을 읽은 후 읽은 내용을 그대로 반복해 옮기기와 그 내용을 응용해 어떤 문제를 푸는 두 가지 과제가 주어졌다. 그 결과 읽은 것을 그대로 옮기는 단순과제에서는 두 그룹 사이에 의미 있는 차이가 발견되지 않았으나 좀 더 복잡한 두 번째 과제에서는 명랑한 기분의 그룹이 훨씬 우수한 처리능력을 과시했다. 위에 박사는 이에 대해 「우리의 지식은 마치 그물과 같은 형태로 기억 속에 기록이 되며 한 개 한 개의 단위지식 (그물의 매듭) 은 서로 복잡하게 연결된다. 그런데 정신활동의 과정에서는 필요한 매듭이 활성화되면서 문제해결에 함께 영향을 끼칠 수 있는 다른 매듭들을 제어 관리하게 된다」고 밝혔다.
기분이 좋을 때는 이러한 제어 관리의 막힘이 없이 문제처리를 위해 개인의 기억 속에 보유한 모든 처리능력을 동원시킬 수 있지만 기분이 나쁜 상태에서는 매듭들의 일부가 우울한 기분을 극복하는데 필요하기 때문에 그만큼 문제해결능력을 떨어지게 된다는 것이다. 즉 명랑한 때는 신경세포를 연결해주는 시냅스에서의 신경전달물질의 유리가 보다 원할하게 이루어져 신경전도가 억제됨이 없이 순조롭게 이루어지나 우울할 때는 시냅스에서의 전도가 더디게 일어난다.
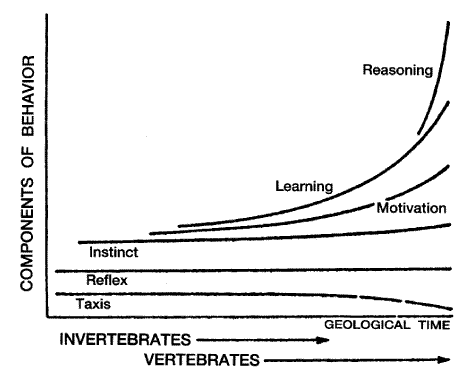
중추신경계가 행동을 조절하는 양상은 개체에 따라 다양하다. 단순반사나 본능은 대다수의 무척추동물이나 하등척추동물의 생활을 지배하나 고등동물에서는 동기 있는 행동 (motivated behavior) 이 지배한다.[그림 1]에는 개체에 따른 여러 가지 형태의 적응행동 (adaptive behavior) 의 발달과정이 도시되어 있다.
[그림 1] 개체에 따른 적응행동의 발달
인간과 고등동물은 과거의 경험을 바탕으로 하여 행동양식을 조금씩 변화시켜 향상 발전시켜 나갈 능력을 가지고 있다. 이를 학습이라 하는데, 학습이란 과거의 경험을 기억하고, 환경에 적응해 나가는 과정이라 하겠다. 학습된 행동을 학습행동 (learned behavior) 이라 하며, 타고날 대부터 갖추어진 행동 (innate behavior) 과 구별한다. 학습은 중추신경계의 가소성 혹은 적응성의 하나라 생각되며, 이는 또 인류 문명과 문화발달의 원동력이 되어 왔으며, 앞으로도 인간의 무한한 발전의 발판이 될 것이다. 다시 말해서 학습은 경험에 의해 야기된 행동의 적응변화 (an adaptive change in behavior caused by experience) 라고 정의할 수 있다. '적응' 이라는 말은 그 종족의 생존이나 동물의 행동에 '어떤 의미' 를 가지고 있다는 말이다. '변화' 라는 말에는 어떤 자극이 가해지기 전과 후에 측정 가능한 차이가 있다는 의미를 내포하고 있으며 이 변화는 전반적인 대사의 항진이나, 피로감, 손상 등을 단순히 반영해서는 안되며 정상 적응 과정도 포함하지 않는다. 즉 변화는 특히 행동의 변화를 나타내야 한다. '행동' 은 개체의 중추신경계에 의한 조절을 받는 것을 의미하지 말초신경계나 말초 감각 운동신경계에 의해 조절을 받지 않는다.
기억이란 어떤 자극(학습)에 대하여 이를 감각하고 이것을 머리에 아로새켜 두었다가, 자극이 없어진 후 그 정보를 다시 상기 할 수 있는 정신기능을 말한다. 인간에게 기억하는 능력이 없었던들 지적 성장 발전은 없었을 것이다. 사고, 판단, 학습도 따져 보면 모두 기억을 바탕으로 한 대뇌기능이다. 기억의 보유시간이 짧고, 곧 상기할 수 있는 기능을 단기기억이라 한다. 이는 비교적 불안정하며, 두부에 외상을 입거나 전격(전기충격) 등으로 의식이 상실되면 기억은 쉽게 소실된다. 기억은 여러 가지 변형을 입어 확고해지고, 두부 외상 혹은 전격에 의해서도 사라지지 않게 된다. 이러한 기억을 장기기억이라 부른다. 정보가 뇌 속에 확고히 고정되어 기억 흔적으로 남기 때문으로 생각된다. 측두엽과 해마를 파괴하면 기억장해가 온다.
MEMORY
Inferred
Processes → Learning → RETENTION → Retrieval
학습과 기억의 신경기전을 이해하는데 있어 중요한 이론적 근거를 제시한 사람은 1940년대의 두 명의 정신학자인 Donald Hebb 와 Jerzy Konorski 이다. 학습과 기억은 신경회로의 변화를 동반해야 한다는 그들의 이론은 Cajal의 이론을 도입한 것이다. 즉 세포가 활발히 작용할 때 시냅스 연결이 더욱더 활발하고 효과적이 된다는 것이다 ([그림 2]) 참고). 단기기억에서는 시냅스흥분성이 일시적으로 증가하고 장기기억에서는 시냅스의 구조적인 변화가 동반될 수 있다는 사실과 유사한 생각을 Konorski는 하였다. 뇌 기능이 세포접촉 (cell assembly) 과 신경회로에 의해 매개된다는 개념은 그 후 널리 인정받게 되었다. 현재 시냅스에서의 가소성 변화 (plastic change) 가 학습과 기억의 중요한 신경기저를 이루고 있다고 믿고 있다.

[그림 2] Cell assembly 와 Hebb synapse (Hebb, 1949)
동시에 반복적으로 활동을 하는 두 개의 세포는 서로 협력하는 경향이 있으며 한 세포에서의 활성의 증가는 다른 세포의 활성을 증가시킨다는 개념을 제시했다. Area 17에 있는 A 와 B세포가 동시에 활성을 가진다. A 세포는 18부위에 있는 많은 세포와 시냅스를 형성한다. C세포는 17부위로 다시 들어와서 시냅스를 이루며, A세포를 흥분시키는 흥분파에 의해 17부위 세포들이 역시 흥분발사를 하게 된다. 또한 C세포는 B세포와 시냅스를 이루며 다시 18부위 세포들을 흥분시킨다. 이들 부위에서 반복적인 큰 흥분파가 연속되므로 해서 AC와 CB 시냅스에 성장 변화가 나타난다는 설명이다.
학습을 평가하는데 조심해야 할 일을 각성 (arousal) 상태의 변화이다. 동물이 학습평가를 받을 때 일반적으로 각성, 주의집중, 동기화부여가 되어 있기 때문에 학습행동 평가에 조심해야 한다. 동물간에 기억의 전이에 관한 실험에서도 투여한 뇌 추출물이 특이하게 학습이나 기억에 영향을 미쳤다기보다, 비특이적으로 각성상태를 변화시켰기 때문에 나온 결과로 해석된다. [표 2]에 표시한 전기생리학적 실험에서도 비슷한 각성상태의 증가로 학습과 기억의 증가가 일어났다고 믿고 있다 (특이한 학습과 기억기전에 영향을 미친 것이 아니라).
[표 2] 학습과 기억기전 연구들
Anatomical 1. Environmintal enrichment leading to bigger brains, 1964 dendrites, and synapses 2. Effects of use on synapses
Biochemical 1. Increased neuronal activity leading to increased RNA 2. Memory transfer between animals 3. Susceptibility to antimetabolite drugs(memory consolidation requires protein synthesis) 4. Cellular biochemistry of ettects of use on synapses
Electrophysiological 1. Resistance to electroconvulsive shock 2. Changes in EEG activity with learning 3. Changes in single neuron activity with learning 4. Cellular electrophysiology of membrane properties at synaptic terminals |
Bennett et al.,
many authors, 1960s to present
Hyden, 1959
McConnell, 1962 Babich et al., Fjerdingstad et al., Reinin et al., 2960s Flexner et al., 1960s
many authors,1960s to present
Duncan, 1949 Adey, 1960; Grastyan, 1966 sheer, 1970 Rowland, 1968
Olds and Olds, 1961 Ranck, 1973 Fuster and Uyeda, 1971 many authors(see Table 29.3) |
|
For references, see Bennett(1977) |
|
행동은 다양한 형태로 나타나기 때문에 여러 가지 다양한 형태의 학습과 기억이 있다는 것은 놀라운 일이 아니다. [표 3] 에는 주요 형태가 표시되어 있다.
[표 3] 학습과 기억의 형태
|
학습형태 |
기억형태 |
|
단순 학습 (simple) 습관화 현상 (Habituation) 감작 현상 (Sensitization) 관계 학습 수동적(전통적) passive(Classical) 능동적 (Operant) 혐오적 (aversion, one-trial) 복합 학습 (Complex) imprinting 잠재학습 (latent) 관찰학습 (vicarious or observational) |
순간기억 (Immediate) 단기기억 (Short-term) 장기기억 (Long-term)
|
신경세포들은 흥분자극에 대하여 다양한 반응을 나타낸다. 일반적으로 감각수용체들은 반복자극으로 반응이 점차 줄어드는 경향이 나타난다. 이러한 반응의 감소를 습관화 (habituation) 라고 한다. 반면 운동신경섬유를 반복자극하면 근육의 반응이 강해지던가 약해진다. 이와 같이 강한 자극에 반사반응이 강화되는 현상을 감작반응 (Sensitization) 이라 한다. 즉, 자극이라는 학습을 반응의 변화는 신경근접합부에서의 신경전달물질의 유리의 차이로 생긴다고 여기고 있다. [표 4]에는 시냅스에서 일어나는 가소성 변화를 이해하기 위해서 실시한 연구들을 나열한 것이다.
[표 4] 학습과 기억기전에 중요한 시냅스의 가소성 변화에 관한 연구들
Posttetanic potentiation at neuromuscular junction Posttetanic potentiation in sympathetic ganglion Posttetanic potentiation in spinal cord Quantal release of transmitter correlated with posttetanic potentiation and depression at the neuromuscular junction Heterosynaptic interactions and presynaptic modulation Voltage dependence of transmitter rlease
Calcium dependence of transmitter release, voltage dependence if calcium channels [Ca2+](intermal) and synaptic plasticity ICa and synaptic plasticity Membrane potential effects of synaptic potential |
Feng, 1941
Larrabee and Bronk, 1947 Lloyd, 1949 del Castillo and Katz,1954
Dudel and Kuffler, 1961
Hagiwara
and Tasaki; Katz and Miledi, 1967
Rahamimoff, 1968 Zucker, 1974 Shinnahara and Tauc; Nicholls and Wallace; Erulkar and Weight, 1970s |
습관화 현상은 일반적인 현상이며, 반응의 강도변화만 있지, 반응의 성격 자체는 변화가 없다. 이런 점에서 어떤 학자들은 습관화 현상이 진짜 학습의 형태가 아니라고 생각하고 있으나 일반적으로는 초보적인 형태의 학습이라고 여기고 있다. 이러한 현상은 Aplysia에서 잘 관찰되고 있다. Eric Kandel 팀은 아가미 (gill) 와 관 (siphon) 에서의 방어적인 제거반사 (Withdrawal reflex) 에서 습관화 현상을 관찰했다. [그림 3-A]에서 보는 바와 같이 물줄기를 아가미의 관에 뿌려주면 이들 속에 있는 접촉 수용체가 자극되어 수축제거반사가 나타나는데 반복 자극하면 수축제거 반사가 감소된다. 이 감소가 수주간지속되기도 한다. 즉 단기와 장기 습관화현상이 나타난다. [그림 3-B] 에서와 같은 전기 생리학적 분석을 해보면, 감각신경세포에서는 흥분곡선이, 운동세포에서는 흥분성 시냅스 후전위 (EPSP) 가 발생된다. 이때 반복자극으로 ESPS 의 진폭이 감소된다. 진폭이 감소되는 것은 신경전달물질의 유리가 적어지기 때문에 일어난다고 생각된다 (시냅스 후막의 반응성은 영향을 받지 않으나). 그러면 무엇이 신경전달물질의 유리를 감소시키는가? Na+채널을 TTX (tetradotoxin) 로 억제시키면 Ca2+에 의해 활동 전압이 나타나는데, 습관화 현상 때는 Ca2+진폭의 감소가 나타난다. 따라서 Ca2+전류가 단기습관화에 중요한 역할을 하는 것을 알 수 있다 ([그림 3-C]). 정상동물에서는 신경말단의 약 40%가 활성 시냅스 부를 가지고 있으나 장기 습관화 때는 약 10%만이 활성 시냅스부를 가지고 있다 ([그림 3-C]). 즉, 장기습관화 현상 때는 각 시냅스에서의 흥분발사양도 적어지지만, 실제 활성 시냅스 수도 줄어든다.

[그림 3] Aplysia 의 아가미와 관에서 나타나는 습관화 현상
A. 습관화 행동
B. 세포 내 기록실험 반복자극으로 나타나는 감각 세포에서의 활동전압의 감소와
운동신경에서의 EPSP의 감소
C. 좌 : 내양 Ca++ 전류의 감소
우 : 장기 습관화 현상 때의 시냅스 말단의 구조적 변화 (Kandal, 1979; Bailay와
Chen, 1983).
감작현상은 강하거나 해로운 자극을 가할 때 반사반응이 항진되는 것을 말한다. 이것은 습관화현상과 반대되는 현상이지만 여러 가지 점에서 다르다. 이 현상은 반사를 야기하는 자극과 다른 자극에 좌우된다.어떤 강한 자극도 일반적인 각성 기전을 활성화 시킨다. 그러므로 감작반응은 일반적인 각성계의 활성을 동반한다. 예를 들면,우리가 시끄러운 소리에 깜짝 놀란다면, 뒤이은 조용한 소리에도 아주 예민해지게 된다. 이러한 감작반응의 세포기전도 Aplysia의 아가미의 제거반사에서 잘 관찰되고 있다. 동물의 피부에 강한 전기 쇽과 같은 해로운 자극을 연속해서 가하면 습관화 현상이 나타나 있는 아가미 제거반사가 증가되어 거의 정상 진폭을 회복하게 된다 ([그림 4]). 이러한 회복현상은 감각 신경말단에서 자극에 의해 유리되는 신경전달 물질의 양이 증가하기 때문에 나타난다. 그 자세한 발생 과정은 Schwartz 와 Kandel 이 다음과 같이 설명하고 있다.
(a) 세로토닌이나, 다른 전달물질 혹은 펩티드가 유리되어 감각 신경말단을 활성화시킨다.
(b) 활성화된 수용체와 아데닐고리화 효소 (adenylate cyclase) 가 접합되어 cAMP 생성한다.
(c) cAMP-의존성 protein kinase의 활성화
(d) K+채널 (S 채널) 의 인산화로 흥분 자극 시 K⁺ 전류의 감소
(e) 흥분파의 증가로 Ca2+유입의 증가와 전달 물질유리의 증가가 나타난다.
따라서 감작현상은 습관화 현상과 반대로 신경전달물질유리의 증가로 인한 시냅스 활성강도의 증가가 나타난다.
장기간의 훈련은 장기간 지속되는 감작현상을 나타낸다. 전자현미경으로 보면 장기 습관화 때 나타나는 활성시냅스부위 감소 대신 활성부가 65%까지 증가될 뿐만 아니라 더 크고 더 복잡해진다 ([그림 4C]). 나아가서 신경말단부가 돋아난다 (sprouting). 이러한 현상은 2차 전달자계가 단기변화를 일으킬 뿐만 아니라, 시냅스부위의 장기적 구조변화를 일으키는데 필요한 단백질 합성도 증가시킨다는 의미이다. 단백질 합성 억제제를 투여하면 단기 감작현상에는 영향을 미치지 않고 장기적 감작현상과 장기적 구조변화가 나타나지 않는다.
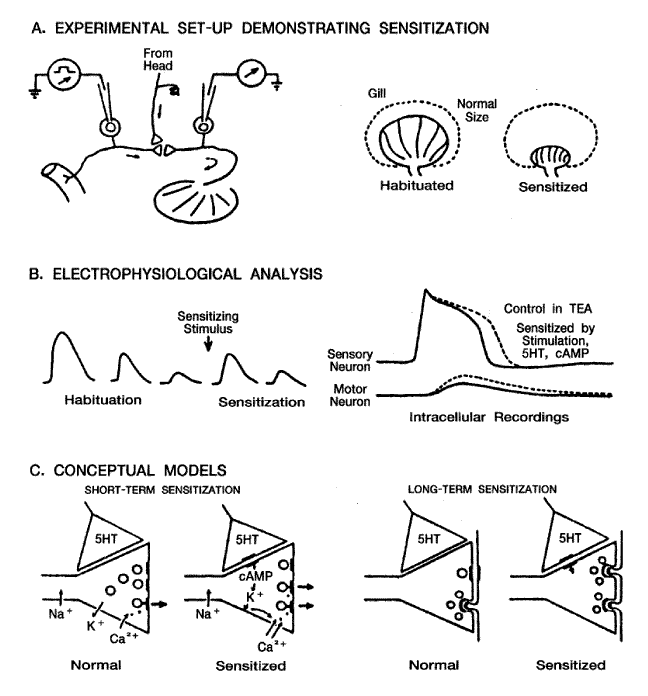
[그림 4] Aplysia에 나타나는 아가미 제거 반사의 감작현상
A. 습관화 현상과 머리로부터의 유해수용체 (nociceptor) 자극에 의한 감작현상
B. 좌 : 습관화 현상과 감작현상시의 아가미 반응의 진폭
우 : 아가미 반사의 감작현상 때 감각 세포 활동전압의 증가와 운동세포 EPSP의
촉진
C. 좌 : 감작시 나타나는 2차 전달자계와 인산화 반응 및 K+전류의 감소와 Ca2+
전류의 증가
우 : 장기 감작현상 때 나타나는 구조적 변화 (Kandel 1979, Shapiro 등 1980,
Bailey와 Chan, 1983 인용)
관계학습에서 동물은 처음자극과 두 번째 자극사이에 관계를 맺어 행동반응을 나타내게 된다. 가장 전형적인 예가 1900년대의 이반 파블로프에 의해 기술된 조건반사 (conditioned reflex) 이다. 이것이 전형적인 조건화 (classical conditioning) 현상이다.
Siphon-gill 반사도 전형적으로 조건화시킬 수 있다. [그림 5]에서 보는 것처럼 강한 전기 쇽 (무조건자극) 을 꼬리에 가하고 덮개에 약한 쇽 (조건 자극) 을 동시에 가하면 운동세포에서의 단일 시냅스 반응이 증가된다 ([그림 5C]). 이런 현상이 나타나는 기전은 감작현상 때와 유사하다. 즉, 조건화 현상 때 Ca2+ 의 유입이 증가하며 Ca2+ 이 calmodulic과 결합하여 cAMP 형성이 증가한다. 이어 cAMP 의존성 protein kinase 가 활성화되어 K+(s) 채널이 인산화된다. 결과로 K+채널이 닫혀서 Ca2+의 세포 내로의 유입이 증가되며 전달물질의 유리가 더 많아져서 시냅스 후 세포반응이 증가된다.
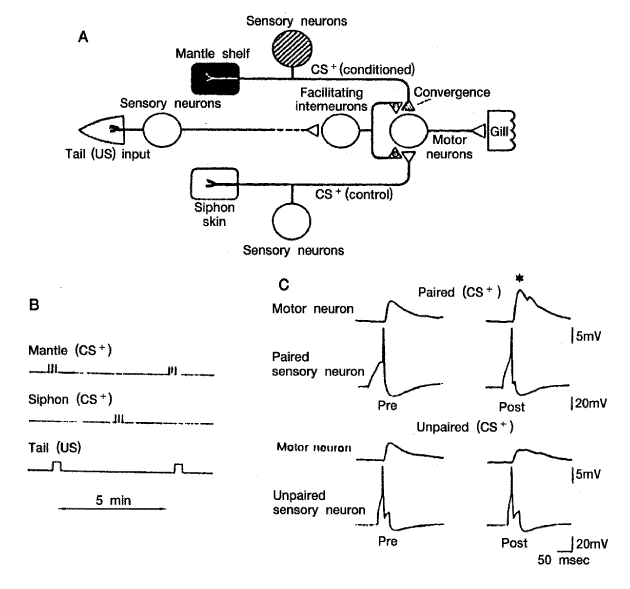
[그림 5] Aplysia에서 나타나는 전형적 조건화 현상의 기전
A. 아가미 반사를 조절하는 신경계, 무조건자극 (US) 으로 강한 전기 쇽을 꼬리의
감각신경에 가하면 언제나 아가미 수축반응이 나타난다. 조건자극 (CS+) 으로 약한
전기 쇽을 덮개의 감각신경에 가하면 그 자체로서는 반응을 일으키지 않는다. 대조의
조건자극 (CS-) 으로 약한 전기 쇽을 siphon 피부의 감각 신경에 가한다.
B. CS⁺를 US와 짝을 맞춘다.
C. 조건화 전 (pre) 과 조건화 1시간 후 (post) 세포 내 기록 검사를 해보면 CS+와
US가 동시에 짝을 맞추어 가하면 운동세포의 EPSP의 강화가 나타나나 CS-는 안 나타난다
(Kandel 과 Schwartz, 1985인용)
1986년 Thompsom 은 기억혼은 뇌저나 척수와 같은 저위 반사 센터에 있는 것이 아님을 관찰했다. 주로 소뇌, 해마, 편도핵과 대뇌피질에 있음이 밝혀졌다.
① 소뇌
모든 종류의 운동을 조절하는 소뇌는 사과를 집어 입에 넣는 것과 같은 여러 종류의 운동을 적절히 조화 연결시켜주는 것 같다. 소뇌가 손상되면 일련의 연결된 복합행동은 못하고 중간에서 행동이 단절된다 (사과를 집어 올리나, 입에 넣지를 못함). McCormick 등 (1982)은 많은 전형적 조건반응이 소뇌에 저장됨을 주장하였다. 토끼 눈에 공기바람을 쏘이면 (무조건 자극) 눈꺼풀이 윙크한다. 이때 공기바람과 함께 소리자극 (조건자극) 을 반복해서 같이 주면 소리자극만으로도 윙크하게 된다. 이후에 토끼의 소뇌 (같은 쪽) 를 아주 소량 제거하면 조건반응은 완전 소실하나 무조건 반응 (공기바람에 윙크하는) 은 그래도 남아있다. 눈꺼풀의 조건반응을 야기하는 기억혼은 심부소뇌핵에 있는 것 같다. 이 중에서 lateral interpositus nucleus 가 중요한 부위이다. 무조건 반사의 주요 구심경로는 inferior olive 이며 주요 원심경로는 red nucleus 이다.

[그림 6] 소뇌는 모든 운동반응, 적어도 전형적인 조건반응의 학습에 필수적인 것 같다.
② 대뇌피질
학습과 기억에 중요하다는 것은 의심의 여지없다. 습관화 (Habituation) 나 전형적 조건화 반응과같은 단순학습은 뇌고위 중추의 기억이 필요치 않는 것 같다.
전형적 조건 반응에서는 동물은 수동적이다. 반면에 능동적 조건 반응에서는 동물은 능동적으로 상자에서 빠져 나온다든지, 지렛대를 누른다든지, 미로를 찾아 나온다든지 하는 문제 해결반응을 보인다. 이 실험방법은 1898년 Edward Thorndike 가 개발한 것이다. 동물은 보통 일을 배우기 전에 실수를 저지르기 때문에 trial-and error 학습이라고도 불려진다. 전형적 (수동적) 조건반응과 능동적 조건반응 모두에서 반응의 강도는 보상 (reward) 이나 벌 (punishment) 의 양에 비례한다. 특히 능동적 조건반응에서는 반응의 강도를 보상을 얻고자 하거나 벌을 피하고자 하는 동물의 자발성의 척도로 사용될 수 있다. 1961년 James 와 Marianne Olds 에 의해 처음으로 관찰되었다. [그림 7] 에 원숭이 대뇌운동피질 세포에서 Eb Fetz 와 Mery Ann Baker (1973) 가 수행한 실험 결과가 도시되어 있다. 기록은 뇌에 고정된 미세전극을 통하여 이루어졌으며 흥분발사율이 일정 값 이상 초과하면 달콤한 과일 주스 및 방울을 상으로 주었다. 훈련기간동안 흥분발사율이 증가하였으며, 점차 강해지고 빈도가 단축되었다. 보상을 제거하면 흥분발사율이 급격히 훈련 전의 측정치로 감소되었다. 피질 신경세포의 흥분 시 바로 뒤이어 근육운동의 흥분이 나타났다 ([그림 7D]).
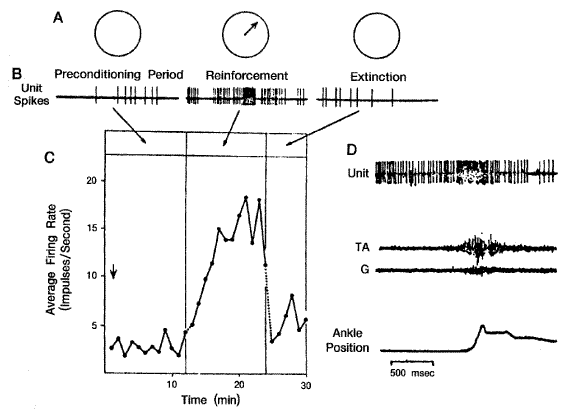
[그림 7] 원숭이 대뇌운동피질에 있는 한 개의 신경세포에서의 능동적 조건반응.
A. 흥분발사율이 미리 정해놓은 일정 값을 초과할 때 다이얼이 점멸하고 보상을
내린다.
B. 흥분발사율의 기록
C. 훈련기간동안 흥분발사율의 증가
D. 근육활성도의 EMG 기록, TA (Tibialis anterior) G (Gastrocuemius) 운동피질신경세포의
흥분발사 시의 족관절 운동
1966년 John Garcia와 Robert Koelling 은 쥐가 어떻게 맛과 질병을 연관시키는 가에 관심을 가지고 다음과 같은 실험을 하였다. 우선 방사선을 쥐에 조사해서 위장관의 손상을 야기했다. 몇 시간 후 쥐는 위장관 질병을 앓게 되었으며 이때 달콤한 물을 주었다. 쥐가 질병에서 회복한 후 다시 달콤한 물을 주었을 때 쥐는 먹으려고 하지 않았다. 즉 단물이 아프게 만들었다고 학습되었기 때문에 단물에 대한 맛을 잃고 거절하게 된 것이다. 이런 학습은 단지 한번만의 경험을 필요로 하기 때문에 One-trial learning 이라고도 불려진다. 무조건 자극 (질병)과 조건자극 (맛) 사이에 연관이 맺어 졌기 때문에 관계 학습 (associative learning) 에 속하는 전형적 조건 반응과는 몇 가지 차이점이 있다. 보통 전형적 조건반응에서는 무조건 반응이 조건반응으로 전이가 되기 위해서는 수많은 시도가 필요하며 무조건자극과 조건자극 가하는 시간간격이 아주 가까워야 연계를 맺을 수 있다. 그러나 혐오학습에서는 한번만의 시도로 충분하며 무조건자극과 조건자극 가하는 시간 간격이 수 시간 이상 경과 후에 나타난다. 무조건 혐오자극으로 사용되는 약물로는 Lithium HCI, 암페타민이나 아포모르핀 같은 향정신병 약물들이 있으나 strychnine 같은 독성물질, 일반적인 스트레스, 수동과 능동조건 반응에서는 효과가 있는 전기 쇽은 무조건 혐오자극으로 효과가 없다. 혐오학습은 미각으로부터 자극이 들어올 때 가장 효과적이지만 혀를 촉각 자극할 때도 나온다. 이러한 혐오학습의 신경경로는 어떻게 되어 있는가? 확실히는 밝혀져 있지는 않지만 주로 감각의 도입은 장관구심 신경을 통해서 이루어지며 제 10번 두개골 신경을 통해 뇌저로 올라가서 고속핵 (Nucleus of solitary tract) 으로 간다. 이 고속핵은 맛과 장관의 구심성 흥분이 모이는 장소일 뿐만 아니라 각성상태를 조절하는 중요한 부위이다 ([그림 8]). 또한 편도핵 (Amygdala) 도 혐오학습에 주요한 부위이다. 이 부위의 손상을 가진 동물은 정상적으로 공간, 위치감각을 가질 수가 없다. 즉 행동의 동기화가 없게 된다. 이 혐오학습의 기전에 대한 가설을 보면 습관화나 감작반응과 비슷하게 채널단백질의 인산화, K+, Ca++ 채널전도의 변화가 일어나서 생긴다고 생각되고 있다.

[그림 8] 포유동물의 혐오학습에 관계되는 신경경로 (인용 : Ashe and Nachman, 1980)
어린 동물이 부모에 집착적인 행동을 나타내는 것을 말한다. 아주 어린 시기의 얼마동안 부모의 신체모양이나 깃털의 색깔과 같은 특별한 자극에 따라가는 집착을 보인다. Konrad Lorenz 의 산책길에 뒤를 따르고 있는 어린 거위의 유명한 그림이 이 예이다. 이러한 행동이 나타나지 않으면, 이후의 성장과 정이 비정상적으로 된다. 노래하는 새가 노래를 배우지 못하고 성장이나 성적 행동이 나타나지 않게 된다. 따라서 Imprinting은 필수적인 학습의 행동이다. 이때도 단백질과 RNA의 합성이 증가되는 것이 관찰되며, 시냅스 수나 표면적은 증가한다.
동물은 미로 같은 실험환경에 노출시켜 달리게 하면 (훈련이나 보상 없이) 그 당시에는 학습의 증거가 나타나지 않으나, 후일 미로에 다시 넣었을 때는 노출 안 시킨 동물보다 더 빨리 능동적인 일을 배우게 된다. 이와 같은 경험을 한 동물은 시냅스 크기도 크고 뇌도 커진다는 실험증거들이 있다.
동물이 다른 동물이 하는 일을 볼 때 더 빨리 그 일을 배우게 된다. 사람에서 아주 중요한 학습이다. 무척추동물이나 하등척추 동물에서는 이런 관찰학습을 나타나지 않는다.
지난 세기동안 일어난 가장 중요한 발견들 중의 하나는 뇌는 많은 요인들에 대해서 끊임없이 변한다는 사실이다. 예를 들면 학습으로 인하여 행동의 변화가 나타날 때 신경세포들 사이의 연결에 기능적 변화가 동반된다. 축삭이나 수상돌기가 변화되는 것과 같은 가벼운 변화에서부터 새로운 신경세포가 형성되는 등의 더 심한 변화가 일어난다 (fish, bird, some mammals). 뇌는 수많은 뉴론 들이 회로를 형성하며, 서로 상이한 회로가 서로 다른 정보를 서로 다른 방법으로 처리한다. 경험에 의해서 뇌 회로의 기능이 변화할 때, 이러한 변화가 "학습" 을 야기한다. 많은 연구자들은 경험에 의존하는 회로 (experience dependent circuit) 가 어떻게, 어디에서 일어나는 지 즉, 기억이 어떻게, 어디에 저장되는지를 연구하고 있다. 바다 달팽이 (sea slug) 는 크고 간단한 신경계를 가지고 있기 때문에 기억연구에 많이 이용되고 있다. 이 실험의 결과, 경험에 의존적인 화학변화는 처음 시냅스에서 일어나며 이러한 변화가 기억의 분자기전을 이루고 있다. 그러나 시냅스는 장기기억을 저장하기에는 안전한 장소가 아니기 때문에 아마 세포의 핵이나 유전정보가 함유되어 있는 염색체에 저장되는 것 같다. 앞으로의 연구 초점은 어떻게 단기기억이 장기기억으로 바뀌는 가를 연구하는 것이다. 즉, 어떻게 세포가 유전자 발현을 조절하는가? 어떻게 유전자 발현과 행동이 서로 영향을 미치는가? 를 연구하는 것이다. 고빈도의 강한 자극을 세포에 반복적으로가하면 전기적 반응이 강화되고 오래 지속된다. 이런 현상을 장기강화 (long-term potentiation) 라고 한다.
해마의 과립세포를 고빈도 (tetanically) 로 수초 자극하는 것을 되풀이하면 정상보다 진폭이 훨씬 커진다 ([그림 9]). 이런 현상을 장기 강화 (long-term potentiation) 라 한다. 이 현상은 신경근 시냅스에서 관찰되는 강직 후 강화 (posttetanic potentiation) 와 비슷하나 훨씬 더 강력하다. Dendate 세포에서는 수일~수주간 지속적인 관찰이 가능하다. 이 LTP 현상에서 시냅스 전 세포에서는 Glutamate 유리의 증가가 나타나고 더 나아가 aplysia 에서와 비슷하게 이온채널과 많은 단백질의 인산화도 관찰되고 있다. 시냅스후 세포에서는 Glutamate의 NMDA 수용체수가 증가하여 ( Ca++-calpain-fodrin) 더 많은 Ca++이 세포 내로 들어온다. LTP는 Ca2+chelator 인 EGTA 에 의해 차단된다.
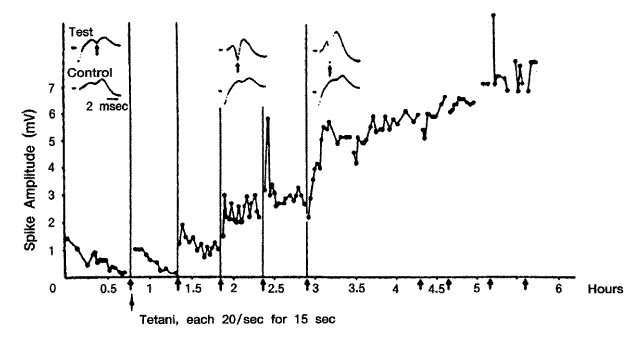
[그림 9] 오래 지속되는 해마 과립세포에서의 장기강화
1초에 20번 전기 자극을 15초간 주는 것을 반복하면 (화살표) 진폭의 상승이 나타난다. 6시간 까지 기록함
이러한 LTP (시냅스 장기 강화) 현상은 척추동물 뇌에서 학습과 기억 기전을 연구하기 위한 가장 좋은 예이다. 장기 강화가 일어나기 위해서는 두 가지 조건이 필요하다
(a) 시냅스를 고빈도로 자극해야 한다.
(b) 이 자극은 어떤 역치 이상의 강도로 해야 한다.
고빈도의 약한 자극은 LTP를 낼 수 없다. 그러므로 LTP를 나타내게 하기 위해서는 상당한 수의 시냅스전 섬유가 활성화되어야 한다 (협동성). LTP를 나타내는 시냅스는 Glutamate 시냅스이며 주로 NMDA 수용체가 관계가 있다. 이 NMDA 수용체 채널이 활성화되면 Na+, K+ ,Ca++ 의 투과성이 증가한다. Glutamate 이외 다른 전잘물질의 작용가능성도 많이 제기되고 있다. 고위 인지기능과 관계 있는 노르에피네프린 도파민, 세로토닌, 아세틸콜린 중에서 아세틸콜린의 중요성이 증가하고 있다.
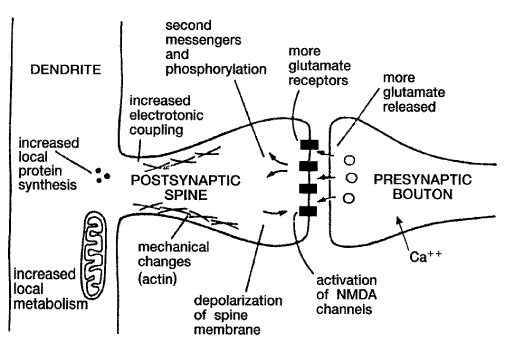
[그림 10] 해마에서 일어나는 LTD의 생성 기전
시냅스전 (presynaptic) 기전 혹은 시냅스 후 기전 (postsynaptic) 에 의해서 장기 강화가 일어나는지는 완전히 밝혀져 있지 않지만 부위에 따라서 한쪽 혹은 양쪽 기전이 모두 작용하고 있는 것 같다. 즉 해마의 CAI 신경세포와 associational-commissural(a-c) 시냅스에서는 시냅스전 기전에 의해 Ca2+의 시냅스 전 세포로의 이동이 증가되고 들어온 Ca2+은 calpain 효소를 활성화시키고 활성화된 calpain (Ca2+activated cytosolic, cysteine protease) 은 fodrin 이라는 막단백질을 분해시켜서 이 속에 감추어져 있던 Glutamate 수용체를 노출시켜 활성화시킨다. 더 많은 수용체가 Glutamate 와 반응함으로써 Ca2+의 이동은 더 증가하게 되고 이차전달자 (second messenger) 계가 활성화되어 뉴론 내의 많은 단백질이 인산화되고 그 결과 단백질 합성이 증가되어 시냅스는 여러 가지 구조적 변화를 겪게 되어 기억이 이 속에 안정하게 고정되는 것 같다. 단백질합성을 억제하면 단기기억은 장애가 나타나지 않으나 장기기억은 장애가 나타난다.
어린 시절에는 calpain 분자와 시냅수후 수용체 이용도가 높기 때문에 어린 시절에는 가소성이 높으나 나이가 들면 calpain 효소가 많이 소모되어 시냅스후막이 더 이상 시냅스전 자극에 반응하지 못한다는 것이다. Lynch는 노인성치매 (알츠하이머병) 에서 이러한 일이 일어날 것이라고 추측했다.
케타민 마취제는 심한 기억상실증을 야기하는데, 이 약이 Ca++ 의 세포 내로의 이동을 심하게 억제한다. 이러한 사실이 위에서 설명한 기억의 분자기전을 잘 증명해 주고 있을 뿐만 아니라, 알츠하이머병과 같은 기억장애질환 수용체의 유전자를 분리하여 어떻게 세포내의 변화가 이러한 유전자들의 발현에 영향을 미치는가를 연구하는 것이다.
기억형성과 기억재생의 약화를 치료할 수 있거나 학습과 기억을 강화시켜 줄 수 있는 약물개발은 '과학의 중요한 도전 (Challenge of Science)' 이 되고 있다. 어떠한 요인들이 기억정보의 일차적이 저장에 관여하며 단기저장을 영구적인 저장으로 바꾸게 하는가에 대해서는 아직 잘 모르고 있다. 일반적으로 신경흥분도와 각성상태를 증가시키는 약물들이 단기 기억과 장기기억의 강화에 도움을 준다.
기억을 항진시키거나 억제시킬 수 있는 약물들을 보면 다음과 같다.
[표 5] 기억에 영향을 미치는 약물들
Impairment of retention Enhancement of retention |
Neuronal excitablity or neural activity Anaesthetics, Hypnotics, Sedatives Central stimulants, low dosage Analeptics, convulsive dosage Analepties, subconvulsive dosage Picrotoxin, subconvulsive dose, Picrotoxin, subconvelsive dose, intrahippocampal systemic Amphetamine, high dose amphetamine, low dose Synaptic transmission and modulation Cholinergic system Scopolamine, Benactyzine, Scopolamine Physostigmine Oxotremorine, Physostigmine Noradrenergic system Phenoxybenzamine Clonidine Reserpine,syrosingopine Dopaminergic system Neuroleptics Apomorphine Serotoninergic system 5-HTP Mianserin, PCPA GABA-system Depakine Peptidergic system Oxytocin Vasopressin Substance P(N.amygdal.) Substance P(median forebrain bundle) Morphine, Naloxone Levorphanol(N.amygdal.) Naloxone(N. amygdat.) Enkephalin ACTH Macrolmolecular metabolism Inhibitors of RNA-synthesis RNA, Uridine, Orotic acid Inhibitors or protein synthesis Mg-Pemoline Galactosamine Amino acids Fucose Other influences Most neurotropic drugs in Piracetam high doses Meclofenoxate Ouabain |
[표 6] 에는 단기기억, 중기기억, 장기기억의 특성들을 요약해 놓았다.
[표 6] 기억의 특성에 관한 요약
Memory phase Formation Duration Regulatory mechanisms |
Short-term memory(STM) Synaptic regulation Operative store for Translocation of electrolytes Conformaional changes of information processing Ca2+, K+ presynaptic, existing membrane proteins during learning postsynaptic (post-stimulatory facilitaion) (post-tetanic potentiation)
Intermediate memory(IM) Synaptosomal regulation Store for ingormation of Release of mediators Phosphorylation of existing short-term significance Mediator-sensitive membrane proteins nucleotide-cyclase Change of transmitter Translocation of electrolytes metabolism Transient store Ca2+-sensitive protein kinases (post-tetanic potentiaion) until long-term memory presynaptic, postsynaptic has been completed
Long-term memory(LTM) Nuclear regulation Release of mediators Phosphorylation of extranucl- Permanent store Mediator-sensitive nucleotide- ear enzymes, change of cyclase activity, Protein synthesis Translocation of electrolytes Glycoprotein formation Ca2+ -sensitive protein kinases Phosphorylation of postsynaptic nucleoproteins Change of gene expression RNA synthesis Protein synthesis Dendritic transport Reconstruction of short-term activated synaptic connections |